Cathepsin-b and cathepsin-l expression levels do not correlate with sensitivity of tumour cells to tnf-α-mediated apoptosis
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT Recently, evidence has been accumulated that besides the caspase proteases, lysosomal cathepsins may play a role in apoptosis induction. This is especially significant as many human
tumour cells express high levels of cathepsins, which might sensitise these cells to specific proapoptotic stimuli mediated by cathepsins. We found that TNF-_α_-mediated DNA fragmentation
in tumour cells was significantly reduced in the presence of E64d and CA074Me, two inhibitors of lysosomal cysteine proteases. Transient transfection of cathepsin-B (Cath-B) and -L (Cath-L)
resulting in expression levels comparable to those found in many tumours did not sensitise tumour cells to TNF-_α_-mediated apoptosis. As lysosomal proteases are thought to be activated by
their release from this organelle into the cytosol, we used the lysosomotropic detergent _N_-dodecyl-imidazole-HCl (NDI-HCl) to disturb lysosomal integrity efficiently and trigger the
release of its proteolytic content into the cytosol. Treatment of HeLa cells with NDI-HCl resulted in cell death, which, however, could also not be influenced by augmented Cath-B or -L
expression levels. Therefore, our data do not support the hypothesis that the high Cath-B or -L expression levels frequently detected in tumour cells might be exploited to target selectively
those tumours for an enhanced cell death effect induced by lysosomotropic agents. SIMILAR CONTENT BEING VIEWED BY OTHERS CASPASE-8 DEFICIENCY INDUCES A SWITCH FROM TLR3 INDUCED APOPTOSIS TO
LYSOSOMAL CELL DEATH IN NEUROBLASTOMA Article Open access 19 May 2021 ALLY OR TRAITOR: THE DUAL ROLE OF P62 IN CASPASE-2 REGULATION Article Open access 14 November 2024 THE CASPASE-2
SUBSTRATE P54NRB EXHIBITS A MULTIFACETED ROLE IN TUMOR CELL DEATH SUSCEPTIBILITY VIA GENE REGULATORY FUNCTIONS Article Open access 20 April 2022 MAIN The expression levels of the lysosomal
papain-like cysteine proteases cathepsin-B (Cath-B) and cathepsin-L (Cath-L) are frequently strongly elevated in tumours of diverse origin. Their secreted forms are thought to contribute to
cancer progression and metastasis either by directly degrading the extracellular matrix or by proteolytically activating other extracellular proteases (Chauhan et al, 1991; Duffy, 1996; Kos
and Lah, 1998; Yan et al, 1998; Sloane, 1990; Turk et al, 2000; Konduri et al, 2001). Therefore, Cath-B and Cath-L are usually regarded as tumour markers correlated with unfavourable
clinical prognosis. On the other hand, in recent years, evidence has been collected that lysosomal proteases and particularly Cath-B might be involved in the mediation of apoptotic cell
death triggered by various stimuli. This hypothesis has been built on observations that, firstly, lysosomal integrity is disturbed during the apoptotic process with concurrent release of the
lysosomal content into the cytosol (Guicciardi et al, 2000; Foghsgaard et al, 2001; Kagedal et al, 2001; Mathiasen et al, 2001; Neuzil et al, 2002). Secondly, cathepsin inhibitors can
reduce the response of cells to some apoptosis inducers (Foghsgaard et al, 2001; Kagedal et al, 2001; Katz et al, 2001; Kiso et al, 2001; Li et al, 2001; Mathiasen et al, 2001). Thirdly,
cells deficient or downregulated in Cath-B are more resistant to TNF-mediated apoptosis (Guicciardi et al, 2000; Foghsgaard et al, 2001). Fourthly, _in vitro_ cathepsins can trigger
cytochrome _c_ release from the mitochondria into the cytosol (Guicciardi et al, 2000; Stoka et al, 2001). This and the fact that tumours frequently contain high levels of cysteine proteases
led to the hypothesis that Cath-B-like proteases may prove useful in selectively targeting tumour cells for apoptosis induction (Foghsgaard et al, 2001). In this report, we show that
TNF-induced cell death in tumour cells could be reduced in the presence of cysteine protease inhibitors E64d and CA074Me, but that overexpression of Cath-B and Cath-L comparable to the
levels detected in tumour cells did not increase the apoptosis sensitivity of those cells to TNF. Even the proapoptotic effect of the lysosomotropic detergent _N_-dodecyl-imidazole, which
directly causes lysosomal instability, was not affected by the intracellular content of Cath-B or Cath-L. Thus, our data do not support the hypothesis that high levels of cathepsin
expression might result in the enhanced response of tumour cells to proapoptotic stimuli. MATERIALS AND METHODS MATERIALS All fine chemicals were obtained from Sigma-Aldrich, unless
otherwise specified. TNF-_α_ was obtained from Biomol, the pan-caspase inhibitor zVAD-fmk was from Enzyme Systems Products, E64d was from Biomol or Peptides International, CA074Me was from
Calbiochem or Peptides International, zFR-AMC was from Enzyme Systems Products or Biomol, _N_-dodecyl-imidazole was from Toronto Research Chemicals Inc., and its hydrochloride salt was
prepared essentially as given in Dubowchik et al (1995). CELL CULTURE HeLa cells and McA-RH7777 cells (kindly provided by Professor Dr GM Kostner, Graz) were cultured in DME medium (Sigma)
supplemented with 2 mM L-glutamine, 1 mM pyruvate, 100 U ml−1 penicillin+100 _μ_g ml−1 streptomycin or 100 _μ_g ml−1 gentamycin and 10% FCS. PC3 cells were kept in RPMI medium (Gibco)
containing the same supplements as described above for the DME medium. Cells were incubated at 37°C at 5% CO2. CONSTRUCTS Coding sequences of hCath-L (Accession M20496), hCath-B (Accession
L16510), and hCath-D (Accession M11233) were obtained by RT–PCR from human cDNA and were cloned into the pcDNA3Δ vector, which was derived from the pcDNA3 vector (Invitrogen) by deletion of
the neomycin resistance region. The obtained cDNA sequences were sequenced and compared to the above given Genbank sequence entries. For stable transfection experiments, the corresponding
constructs were subcloned into the complete pcDNA3 vector containing the neomycin resistance cassette. TRANSFECTIONS HeLa cells and McA RH 7777 cells were transfected using the Effectene
transfection reagent (Qiagen). For this, 0.2 _μ_g plasmid DNA together with 50 ng pEGFP (Clontech) were diluted in 100 _μ_l of EC-Buffer, 1.6 _μ_l enhancer was added, and after 5 min of
incubation at RT 2 _μ_l of the effectene transfection reagent was added, followed by 15 min of incubation at RT. The transfection mix was given to HeLa cells in a six-well plate, which prior
to that had been washed with PBS and then 2 ml of fresh DME medium (10% FCS) had been added. The medium was changed 10 h post-transfection. This protocol could be up- and downscaled for
different culture formats. The transfection efficiency could be controlled by fluorescence of the co-transfected pEGFP and was determined by FACS analysis. PI-FACS ANALYSIS Apoptosis
detection by sub-G1 DNA content was performed by propidium iodide FACS analysis as described (Mund et al, 2003). MTT ASSAY Cells were transfected in 10-cm dishes as described; 24 h later
cells were harvested and reseeded in equal numbers (approximately 7500 cells/well) in the wells of 96-well plates. At 24 h after reseeding, cells were treated as indicated in a total volume
of 100 _μ_l DME medium. At the time of analysis, MTT assays were essentially performed according to Mosmann (1983). ‘Loss of viability, %’ was calculated using the following equation:
(_U_−_S_)/_U_*100, where _U_ is the absorption value of the untreated control and _S_ is the absorption value of sample. IMMUNOBLOTTING For detecting protein expression, cells were harvested
by trypsinisation, washed with PBS, and lysed in Triton-X buffer (50 mM Tris-HCl pH 8.0, 150 mM NaCl, 1% Triton-X 100) 10 min on ice. After centrifugation at 14 000 G in a microfuge,
supernatants were obtained as cytoplasmic extracts, which were quantified for protein content using the Bradford reagent (BioRad). Western blotting was performed as described (Bauer et al,
1999) using either monoclonal anti-Cath-L antibody (Transduction Laboratories) or monoclonal anti-Cath-B antibody (Ab-2, Oncogene Research Products). CATHEPSIN ENZYMATIC ASSAY Cells were
left untreated or were transfected with the pcDNA3Δ expression vector containing the ORFs of Cath-L, or Cath-B, and at the time of analysis the cathepsin activity assay was performed using
zFR-AMC (Enzyme Systems Products) as a substrate, applying a protocol derived from Kamboj et al (1993). AMC fluorescence was measured with a Fluoroskan Ascent FL fluorecence plate reader
(Labsystems) at a wavelength of 380 nm for excitation and 450 nm for emission. For measuring Michaelis–Menten kinetics, the assay was performed as described above in determining the initial
turnover rates of the zFR-AMC substrate at various concentrations (50, 100, 200, 400, and 600 _μ_ M) in the presence of 0.5 _μ_g protein extract from Cath-B overexpressing HeLa cells.
CELL-FREE ACTIVATION OF CASPASES FOR INHIBITOR EXPERIMENTS WITH E64D Cytoplasmic extracts were prepared and activated by the addition of cytochrome _c_ and dATP in the absence or presence of
various concentrations of E64d, and were analysed by a caspase enzymatic assay essentially as described (Gewies et al, 2000). IMMUNOCYTOCHEMISTRY Cells were seeded on cover glasses, and 24
h later were incubated for 16 h in the absence or presence of NDI-HCl, CHX, or TNF/CHX. Immunostaining was carried out as described (Mathiasen et al, 2001) using an anti-Cath-L mAb (1 : 100,
Transduction Laboratories) and an anti-mouse FITC-conjugated polyclonal Ab (1 : 500, Pharmingen) as the primary or secondary antibody, respectively. Fixed and stained cells were covered
with an embedding buffer (0.5 M Tris-HCl pH 8.2, 0.02 g ml−1 DTT, 0.04 g ml−1 polyvinyl alcohol, 40% glycerol), fluorescence was observed under a fluorescence microscope (Axioscop 2, Zeiss),
and pictures were taken with a digital camera (Visitron Systems, Puchheim) using the IPLab imaging software (Spectra Services, NY, USA). RESULTS CYSTEINE PROTEASE INHIBITORS E64D AND
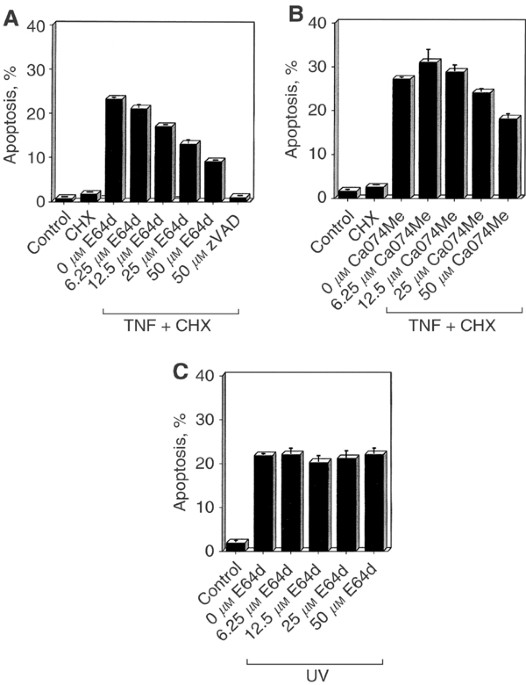
CA074ME SUPPRESS TNF-MEDIATED APOPTOSIS First, we asked whether lysosomal cysteine proteases might be involved in apoptosis induced by TNF in the cervical carcinoma cell line HeLa. Cell
death was assessed by DNA fragmentation in HeLa cells cotreated with TNF and cycloheximide (CHX). The pan-caspase inhibitor zVAD-fmk could completely prevent the demise of the cells,
indicating an apoptotic mode of cell death (Figure 1A). This cell death could also be inhibited in the presence of the broad-spectrum cysteine protease inhibitor E64d in a dose-dependent
manner, albeit not as efficiently as with zVAD-fmk (Figure 1A). We also used CA074Me, a described specific Cath-B inhibitor (Buttle et al, 1992), and likewise detected a decrease of
TNF-induced DNA fragmentation (Figure 2B). Testing the prostatic carcinoma cell line PC3 as a further cell system, we could confirm the inhibitory effect of E64d, CA074Me, and zFA-FMK, a
further Cath-B inhibitor (Rasnick, 1985), on TNF-induced DNA fragmentation (data not shown). To address the question of how general the observed inhibitory effect of E64d on apoptotic DNA
fragmentation is, we tested its impact on UV-mediated cell death in the HeLa cell system. As shown in Figure 1C, E64d did not inhibit apoptosis induced by UV irradiation under the conditions
used. The observation that the Cath-B inhibitor CA074Me is able to reduce TNF-induced apoptosis in HeLa cells (Figure 1B) suggested an involvement of Cath-B in this cell death process.
Consequently, we wanted to prove the specificity of CA074Me on Cath-B activity. To this end, we transiently overexpressed hCath-B or hCath-L in HeLa cells for subsequent inhibitor studies.
Immunoblot analysis demonstrated the efficient overexpression of these two cathepsins, whereas endogenous levels of Cath-L and Cath-B in comparison were quite low (Figure 2A). We also
measured cathepsin activity in extracts of Cath-B- or Cath-L-transfected cells, which were grown for 3 h in the presence or absence of 25 _μ_ M E64d or 25 _μ_ M CA074Me, respectively.
Concentrations in the range of 25 _μ_ M had been used in recent reports (Foghsgaard et al, 2001; Varghese et al, 2001) and were necessary to elicit an inhibitory effect (Figure 1). As
expected, E64d could strongly inhibit both hCath-L and hCath-B enzymatic activity (Figure 2B). Unexpectedly, the supposedly specific Cath-B inhibitor CA074Me not only inhibited the enzymatic
activity of the overexpressed hCath-B but also that of the overexpressed hCath-L. However, this observation is in line with a recent report (Montaser et al, 2002), which revealed that
before cleavage by cellular esterases the methyl ester CA074Me cannot be regarded as a specific Cath-B inhibitor and also affects other lysosomal cysteine proteases such as Cath-L. We also
tested the effect of E64d on caspase enzymatic activity _in vitro_: caspase activity induced by the addition of cytochrome _c_ and dATP to cell extracts in a cell-free system was not
significantly reduced in the presence of 50 _μ_ M E64d in the extract (data not shown), therefore excluding the possibility that the effect of E64d on TNF-induced apoptosis is due to the
concurrent inhibition of caspases. Thus, Cath-B and also related lysosomal cysteine proteases such as Cath-L should be responsible for mediating the E64d- and CA074Me-sensitive TNF signal.
As shown in Figure 2, the overexpression of hCath-B and hCath-L could be obtained by transient transfection of HeLa cells in terms of both protein levels detected in immunoblots (Figure 2A)
as well as cathepsin activities (Figure 2B). In these experiments, we usually observed an increase of cathepsin activity by at least 6- to 10-fold for hCath-L or by 10- to 20-fold for
hCath-B when compared to endogenous cathepsin activity. This extent of increased enzymatic activity is comparable to the situation found in malignant human tumour cells for which elevated
cathepsin-expression and -activity levels have been described (Chauhan et al, 1991; Lah and Kos, 1998; Yan et al, 1998). TRANSIENT OVEREXPRESSION OF CATH-D BUT NOT OF CATH-L AND CATH-B
INDUCES APOPTOSIS IN HELA CELLS Given the possible involvement of Cath-B-like cysteine proteases in TNF-mediated apoptosis in HeLa cells (Figure 1), we decided to investigate the influence
of cathepsin expression levels on the sensitivity of HeLa cells to the TNF cell death signal. Especially, Cath-B has been suggested to play an essential role in TNF-mediated apoptosis (Leist
and Jaattela, 2001), and has been proposed to represent a therapeutical target in cancer therapy of tumours with elevated levels of cysteine proteases (Foghsgaard et al, 2001). We first
examined the effect of increased cathepsin expression on the viability of HeLa cells (Figure 3). We observed cell death in terms of morphology, DNA fragmentation, and caspase-3 enzymatic
activity, when an hCath-D expression vector was transiently transfected into HeLa cells (Figures 3A and B). The ectopic overexpression of Cath-D in HeLa cells was previously shown to induce
cell death dominantly (Deiss et al, 1996). As an additional positive control, we overexpressed the cysteine protease hUBP41, a member of the family of ubiquitin-specific proteases (USPs),
which we recently found to possess dominant apoptosis-inducing activity (Gewies and Grimm, 2003). In contrast, the overexpression of hCath-L and hCath-B did not result in significant cell
death, even though in the case of hCath-L a slight increase in DNA fragmentation values and caspase-3 activity could be detected (Figures 3A and B), concomitant with an increased number of
detached cells (not shown). When we determined the cathepsin activity, we observed an approximately 20-fold increase in these experiments (Figure 3C). AUGMENTED LEVELS OF HCATH-L OR HCATH-B
FAIL TO SENSITISE HELA OR MCA RH 7777 CELLS TO TNF-INDUCED CELL DEATH With our results indicating that cathepsins participate in mediating TNF-induced apoptosis (Figure 1), we speculated
that elevated cathepsin expression levels might influence the sensitivity of HeLa cells to the apoptosis-inducing signal triggered by TNF. As shown in Figure 4A, HeLa cells transiently
transfected with hCath-L or hCath-B did not show any increased apoptosis when compared to control transfected cells. We also generated HeLa cell pools stably overexpressing hCath-L and
hCath-B at levels of two- or four-fold enzymatic activity, respectively, compared to control cells. Also, those HeLa cells did not exhibit an enhanced response to TNF-induced apoptosis
(Figure 4B). Since Cath-B has been reported to play a major role in the apoptosis of hepatocytes (Roberts et al, 1999; Guicciardi et al, 2000, ), we also transiently overexpressed hCath-L or
hCath-B in the rat hepatoma cell line McA RH 7777. The treatment of these McA RH 7777 cells with TNF did not yield any significant differences in apoptosis levels between control- and
cathepsin-transfected cells (Figure 4C). THE LYSOSOMOTROPIC DETERGENT NDI-HCL CAUSES CELL DEATH THAT IS NOT ENHANCED IN HELA CELLS OVEREXPRESSING HCATH-L OR HCATH-B The mode of action of
lysosomal proteases for apoptosis is assumed to involve the disintegration of lysosomes and the release of their content into the cytosol. Subsequently, lysosomal proteases cleave and
activate proapoptotic factors such as the Bcl-2 family member Bid, which leads to the release of cytochrome _c_ from the mitochondria and to the activation of caspases (Guicciardi et al,
2000; Turk et al, 2002). Consequently, we wanted to investigate whether the integrity of lysosomes in TNF-treated HeLa cells is disturbed so that elevated cathepsin levels within the
lysosomes caused by overexpression can be expected to result in higher amounts of cathepsins in the cytosol. As a positive control, we treated HeLa cells with the lysosomotropic detergent
_N_-dodecyl-imidazole hydrochloride (NDI-HCl), which accumulates in lysosomes and eventually damages the lysosomal membrane so that the lysosomal content is released into the cytosol
(Dubowchik et al, 1995). Anti-Cath-L immunofluorescent staining of untreated control HeLa cells generated the expected spot-like lysosomal distribution pattern of the endogenous Cath-L
(Figure 5A). The treatment of HeLa cells with TNF resulted in a considerable number of cells with a decrease of staining intensity of discrete lysosomal spots and an apparent partial
redistribution of Cath-L from those spot-like structures to a more evenly distributed pattern as shown in Figure 5D. The application of NDI-HCl to HeLa cells produced a uniform staining of
Cath-L within still intact cells, indicating extensive lysosomal rupture and release of its content into the cytosol (Figure 5B). We observed that incubation with NDI-HCl led to
morphological changes resembling apoptosis. Subsequent FACS analysis confirmed apoptosis induced by NDI-HCl (not shown). We then asked whether this cell death can be enhanced by
overexpression of hCath-L or hCath-B. For these measurements, we used the MTT assay in order to also include nonapoptotic cell death. As shown in Figure 6, expression levels of hCath-L or
hCath-B did not have any significant effect on NDI-HCl-induced cell death in HeLa cells as determined by the MTT assays. Apoptosis quantification by FACS analysis also did not reveal a
differential cell death induction (not shown). Cell death by NDI-HCl could be partially suppressed in the presence of the pan-caspase inhibitor zVAD-fmk, indicating caspase-dependent
mechanisms. Interestingly, this type of cell death could not be inhibited in the presence of the broad specificity cysteine protease inhibitor E64d (Figure 6). DISCUSSION Caspases are
thought to build the central proteolytic network involved in the execution of apoptotic signalling pathways induced by virtually all kinds of death stimuli (Earnshaw et al, 1999; Nagata,
1999; Robertson and Orrenius, 2000). In recent years, it has been recognised that besides caspases other proteases such as cathepsins, calpains, and the proteasome also may play a role as
cofactors in mediating several cell death stimuli, for example, Cath-B in TNF-induced apoptosis (Leist and Jaattela, 2001). Cath-B has been reported to be released from the lysosomes to the
cytosol in response to TNF (Guicciardi et al, 2000; Foghsgaard et al, 2001) and to contribute to apoptotic downstream events such as cytochrome _c_ release and the activation of executioner
caspases (Guicciardi et al, 2000; Stoka et al, 2001). These observations suggest a possible role for cathepsins as positive effectors of cell death pathways. This and the fact that tumours
frequently contain high levels of cysteine proteases led to the hypothesis that Cath-B-like proteases may prove useful in selectively targeting tumour cells for apoptosis induction
(Foghsgaard et al, 2001). The central objective of this study was to test the hypothesis of whether increased expression levels of Cath-L or Cath-B would enhance the sensitivity of tumour
cells to TNF-mediated apoptosis. We found this not to be the case, even though inhibitor studies provided evidence for an influence of cathepsins in the TNF signalling pathway: apoptotic DNA
fragmentation induced by cotreatment of HeLa and PC3 cells with TNF and CHX could be significantly suppressed by the cathepsin inhibitors E64d and CA074Me in a dose-dependent manner
(Figures 1A and B), which is in line with previous reports (Guicciardi et al, 2000; Foghsgaard et al, 2001). Interestingly, apoptosis induced by irradiating the cells with UV light could not
be diminished in the presence of E64d (Figure 1C). Thus, in HeLa cells, cathepsins appear to contribute to the extrinsic receptor-mediated TNF cell death pathway, but not to the intrinsic
apoptosis signalling pathways induced by UV. The inhibitor CA074Me is usually regarded to be highly selective for Cath-B, and thus one could conclude that Cath-B is the noncaspase cysteine
protease contributing to TNF-mediated apoptosis in the HeLa cell system. When we tried to verify the specific inhibitory activity of CA074Me, we realised that CA074Me not only completely
blocked hCath-B enzymatic activity but also blocked hCath-L activity to almost the same extent (Figure 2B). Our observation was confirmed by a recent publication, also demonstrating that
CA074Me is not a selective inhibitor of Cath-B (Montaser et al, 2002). Therefore, we cannot rule out the fact that besides Cath-B, CA074Me also inhibits other cysteine proteases such as
Cath-L, and by this exerts its inhibitory effect. There are several reports about strongly augmented expression levels of Cath-L and Cath-B detected in tumour cells. Frequently, the increase
in cathepsin expression and activity correlated with malignant progression and metastasis possibly by direct degradation of the extracellular matrix or by activation of other proteases such
as the urokinase-type plasminogen activator (Turk et al, 2000). Consequently, cathepsins are regarded to be tumour markers with their increased expression levels correlating with poor
prognosis (Yan et al, 1998; Sloane, 1990; Chauhan et al, 1991; Sivaparvathi et al, 1995; Duffy, 1996; Lah and Kos, 1998). The tumour-promoting effect of cathepsins is in contrast to the
observation that cathepsins such as Cath-B can mediate proapoptotic signals triggered by TNF (Foghsgaard et al, 2001; Guicciardi et al, 2001; Mathiasen et al, 2001), bile salts (Roberts et
al, 1999), sphingosine (Kagedal et al, 2001), activation of the B-cell receptor (Katz et al, 2001), and L-2,5-dihydrophenylalanine (Kiso et al, 2001). Consequently, we wanted to address the
question of whether this contrast might prove useful in developing a cancer therapy selectively targeting tumour cells for cell death, which express high levels of cathepsins (Foghsgaard et
al, 2001). Even though this concept appears attractive, up to now no studies have been presented investigating a possible correlation between elevated cellular cathepsin levels and
sensitivity to apoptotic stimuli. Since our inhibitor studies indicated a contribution of Cath-B-like cysteine proteases in TNF-induced apoptosis in HeLa cells (Figure 1), we decided to
investigate whether the sensitivity of HeLa cells to the TNF signal is enhanced by increased levels of hCath-L or hCath-B. We found that the overexpression of hCath-L or hCath-B by itself
does not result in overt apoptosis induction in HeLa cells, whereas overexpression of hCath-D led to apoptosis as could be expected from a previous report (Deiss et al, 1996). In these
transient transfection experiments, we usually obtained between 40 and 60% transfection efficiency as judged by GFP cotransfection and an increase of cathepsin activity of at least 6- to
10-fold for hCath-L and 10- to 20-fold in case of hCath-B. This is comparable to the activities frequently detected in tumour tissues (Chauhan et al, 1991; Lah and Kos, 1998; Yan et al,
1998). Consequently, under these experimental conditions we expected to be able to uncover any increase in sensitivity correlated with cathepsin overexpression. However, we did not detect a
significant increase in sensitivity to TNF-induced apoptosis in HeLa cells transiently transfected with hCath-L or hCath-B (Figure 4A). Secondly, we generated HeLa cell pools stably
overexpressing hCath-L or hCath-B, respectively. In this case, we could assume that the majority of cells express the corresponding cathepsin, although at a lower level than in transient
expression experiments. Those stable transfected pool clones also did not show any augmented sensitivity to the TNF death signal (Figure 4B). Thirdly, we used the rat hepatoma cell line McA
RH 7777 for transient expression experiments, which also did not indicate any change in sensitivity to TNF upon cathepsin overexpression (Figure 5C). We chose this additional cell line since
hepatocytes have been described to be influenced by the proapoptotic activity of Cath-B (Roberts et al, 1999; Guicciardi et al, 2000, 2001). The release of lysosomal cathepsins into the
cytosol is thought to be decisive for their apoptosis induction. As shown in Figure 5, lysosomal integrity is apparently disturbed when HeLa cells are treated with TNF-_α_, as indicated by
the partial redistribution of endogenous Cath-L from a lysosomal spot-like pattern to a more diffuse staining. Thus, increased expression levels of cathepsins should also be expected to
result in higher amounts of cathepsin activity in the cytoplasm in response to TNF. How can it then be explained that we observe an inhibitory effect of TNF-induced apoptosis by the
cathepsin inhibitors E64d and CA074Me (Figure 1), but do not see any effect upon cathepsin overexpression (Figures 4 and 5)? Firstly, it cannot be excluded that cathepsin-like proteases
other than Cath-L or Cath-B are mainly responsible for the contribution to TNF-_α_ apoptotic pathways, since the inhibitors E64d and apparently also CA074Me are not specific inhibitors of
distinct cathepsins (Figure 2; Montaser et al 2002). Secondly, it is also possible that the endgenous basal levels of lysosomal Cath-L or Cath-B activity are sufficient to provide the
lysosomal stimulus contributing to the proapoptotic signalling cascade for cell death execution. In this case, downstream or upstream factors other than Cath-L or Cath-B levels could be rate
limiting in the execution process of apoptosis, and an increase in Cath-L or Cath-B levels would not further accelerate or enhance the signal from the TNF receptor. Supporting this, we
obtained the linear relationship of a Lineweaver–Burke plot using overexpressed Cath-B and the test substrate zFR-AMC (data not shown). This indicates that cathepsin proteolysis obeys the
Michaelis–Menten kinetics. Consequently, an _x_-fold increase in Cath-B levels corresponds to an _x_-fold increase of substrate turnover. Therefore, overexpression of Cath-B in our
transfection experiments should correlate with an increased turnover of endogeneous Cath-B substrates. Both possible explanations described above could be valid and would be in line with our
observation that overexpression of hCath-L or hCath-B does not show any death-enhancing effect in NDI-HCl-induced cell death (Figure 6), even though it can be assumed in this case that
treatment with NDI-HCl efficiently released the overexpressed cathepsins from the lysosomal compartments to the cytosol (Figure 5). We must point out that NDI-HCl-induced death behaves
differently from TNF-mediated apoptosis, since NDI-HCl results in cell death that cannot be reduced by E64d, and zVAD-fmk can only partially block cell death (Figure 6), presumably due to
the extreme extent of lysosomal rupture caused by this lysosomotropic detergent (Figure 5B) leading to both a caspase-dependent apoptotic mode and a caspase-independent mode of cell death.
In conclusion, Cath-B-like cysteine proteases seem to contribute to certain aspects of apoptotic signalling pathways that involve the disruption of lysosomes. Whereas the possibility
appeared attractive to exploit the release of cathepsins in specifically targeting tumour cells with elevated cathepsin expression levels for cell death induction, our overexpression
experiments do not support this hypothesis. This is, to our knowledge, the first report directly investigating a possible correlation between cathepsin expression levels and the sensitivity
to an apoptotic stimulus, and further studies could extend these findings to other cell systems and additional cell death stimuli. ACCESSION CODES ACCESSIONS GENBANK/EMBL/DDBJ * L16510 *
M11233 * M20496 CHANGE HISTORY * _ 16 NOVEMBER 2011 This paper was modified 12 months after initial publication to switch to Creative Commons licence terms, as noted at publication _
REFERENCES * Bauer MK, Schubert A, Rocks O, Grimm S (1999) Adenine nucleotide translocase-1, a component of the permeability transition pore, can dominantly induce apoptosis. _J Cell Biol_
147: 1493–1502 Article CAS PubMed PubMed Central Google Scholar * Buttle DJ, Murata M, Knight CG, Barrett AJ (1992) CA074 methyl ester: a proinhibitor for intracellular cathepsin B.
_Arch Biochem Biophys_ 299: 377–380 Article CAS PubMed Google Scholar * Chauhan SS, Goldstein LJ, Gottesman MM (1991) Expression of cathepsin L in human tumors. _Cancer Res_ 51:
1478–1481 CAS PubMed Google Scholar * Deiss LP, Galinka H, Berissi H, Cohen O, Kimchi A (1996) Cathepsin D protease mediates programmed cell death induced by interferon-gamma, Fas/APO-1
and TNF-alpha. _Embo J_ 15: 3861–3870 Article CAS PubMed PubMed Central Google Scholar * Dubowchik GM, Gawlak SL, Firestone RA (1995) The _in vitro_ effects of three lysosomotropic
detergents against three human tumor cell lines. _Bioorganic Med Chem Lett_ 5: 893–898 Article CAS Google Scholar * Duffy MJ (1996) Proteases as prognostic markers in cancer. _Clin Cancer
Res_ 2: 613–618 CAS PubMed Google Scholar * Earnshaw WC, Martins LM, Kaufmann SH (1999) Mammalian caspases: structure, activation, substrates, and functions during apoptosis. _Annu Rev
Biochem_ 68: 383–424 Article CAS PubMed Google Scholar * Foghsgaard L, Wissing D, Mauch D, Lademann U, Bastholm L, Boes M, Elling F, Leist M, Jaattela M (2001) Cathepsin B acts as a
dominant execution protease in tumor cell apoptosis induced by tumor necrosis factor. _J Cell Biol_ 153: 999–1010 Article CAS PubMed PubMed Central Google Scholar * Gewies A, Grimm S
(2003) UBP41 is a proapoptotic ubiquitin-specific protease. _Cancer Res_ 63: 682–688 CAS PubMed Google Scholar * Gewies A, Rokhlin OW, Cohen MB (2000) Cytochrome _c_ is involved in
Fas-mediated apoptosis of prostatic carcinoma cell lines. _Cancer Res_ 60: 2163–2168 CAS PubMed Google Scholar * Guicciardi ME, Deussing J, Miyoshi H, Bronk SF, Svingen PA, Peters C,
Kaufmann SH, Gores GJ (2000) Cathepsin B contributes to TNF-alpha-mediated hepatocyte apoptosis by promoting mitochondrial release of cytochrome _c_. _J Clin Invest_ 106: 1127–1137 Article
CAS PubMed PubMed Central Google Scholar * Guicciardi ME, Miyoshi H, Bronk SF, Gores GJ (2001) Cathepsin B knockout mice are resistant to tumor necrosis factor-alpha-mediated hepatocyte
apoptosis and liver injury: implications for therapeutic applications. _Am J Pathol_ 159: 2045–2054 Article CAS PubMed PubMed Central Google Scholar * Kagedal K, Zhao M, Svensson I,
Brunk UT (2001) Sphingosine-induced apoptosis is dependent on lysosomal proteases. _Biochem J_ 359: 335–343 Article CAS PubMed PubMed Central Google Scholar * Kamboj RC, Pal S, Raghav
N, Singh H (1993) A selective colorimetric assay for cathepsin L using Z-Phe-Arg-4-methoxy-beta-naphthylamide. _Biochimie_ 75: 873–878 Article CAS PubMed Google Scholar * Katz E, Deehan
MR, Seatter S, Lord C, Sturrock RD, Harnett MM (2001) B cell receptor-stimulated mitochondrial phospholipase A2 activation and resultant disruption of mitochondrial membrane potential
correlate with the induction of apoptosis in WEHI-231 B cells. _J Immunol_ 166: 137–147 Article CAS PubMed Google Scholar * Kiso T, Usuki Y, Ping X, Fujita K, Taniguchi M (2001)
L-2,5-dihydro-phenylalanine, an inducer of cathepsin-dependent apoptosis in human promyelocytic leukemia cells (HL-60). _J Antibiot (Tokyo)_ 54: 810–817 Article CAS Google Scholar *
Konduri S, Lakka SS, Tasiou A, Yanamandra N, Gondi CS, Dinh DH, Olivero WC, Gujrati M, Rao JS (2001) Elevated levels of cathepsin B in human glioblastoma cell lines. _Int J Oncol_ 19:
519–524 CAS PubMed Google Scholar * Kos J, Lah TT (1998) Cysteine proteinases and their endogenous inhibitors: target proteins for prognosis, diagnosis and therapy in cancer (review).
_Oncol Rep_ 5: 1349–1361 CAS PubMed Google Scholar * Lah TT, Kos J (1998) Cysteine proteinases in cancer progression and their clinical relevance for prognosis. _Biol Chem_ 379: 125–130
CAS PubMed Google Scholar * Leist M, Jaattela M (2001) Triggering of apoptosis by cathepsins. _Cell Death Differ_ 8: 324–326 Article CAS PubMed Google Scholar * Li W, Dalen H, Eaton
JW, Yuan XM (2001) Apoptotic death of inflammatory cells in human atheroma. _Arterioscler Thromb Vasc Biol_ 21: 1124–1130 Article CAS PubMed Google Scholar * Mathiasen IS, Hansen CM,
Foghsgaard L, Jaattela M (2001) Sensitization to TNF-induced apoptosis by 1,25-dihydroxy vitamin D(3) involves up-regulation of the TNF receptor 1 and cathepsin B. _Int J Cancer_ 93: 224–231
Article CAS PubMed Google Scholar * Montaser M, Lalmanach G, Mach L (2002) CA-074, but not its methyl ester CA-074Me, is a selective inhibitor of cathepsin B within living cells. _Biol
Chem_ 383: 1305–1308 Article CAS PubMed Google Scholar * Mosmann T (1983) Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays.
_J Immunol Methods_ 65: 55–63 CAS PubMed Google Scholar * Mund T, Gewies A, Schoenfeld N, Bauer MKA, Grimm S (2003) Spike, a novel BH3-only protein, regulates apoptosis at the
endoplasmic reticulum. _FASEB J_ 17: 696–698 Article CAS PubMed Google Scholar * Nagata S (1999) Fas ligand-induced apoptosis. _Annu Rev Genet_ 33: 29–55 Article CAS PubMed Google
Scholar * Neuzil J, Zhao M, Ostermann G, Sticha M, Gellert N, Weber C, Eaton JW, Brunk UT (2002) Alpha-tocopheryl succinate, an agent with _in vivo_ anti-tumour activity, induces apoptosis
by causing lysosomal instability. _Biochem J_ 362: 709–715 Article CAS PubMed PubMed Central Google Scholar * Rasnick D (1985) Synthesis of peptide fluoromethyl ketones and the
inhibition of human cathepsin B. _Anal Biochem_ 149: 461–465 Article CAS PubMed Google Scholar * Roberts LR, Adjei PN, Gores GJ (1999) Cathepsins as effector proteases in hepatocyte
apoptosis. _Cell Biochem Biophys_ 30: 71–88 Article CAS PubMed Google Scholar * Robertson JD, Orrenius S (2000) Molecular mechanisms of apoptosis induced by cytotoxic chemicals. _Crit
Rev Toxicol_ 30: 609–627 Article CAS PubMed Google Scholar * Sivaparvathi M, Sawaya R, Wang SW, Rayford A, Yamamoto M, Liotta LA, Nicolson GL, Rao JS (1995) Overexpression and
localization of cathepsin B during the progression of human gliomas. _Clin Exp Metastasis_ 13: 49–56 Article CAS PubMed Google Scholar * Sloane BF (1990) Cathepsin B and cystatins:
evidence for a role in cancer progression. _Semin Cancer Biol_ 1: 137–152 CAS PubMed Google Scholar * Stoka V, Turk B, Schendel SL, Kim TH, Cirman T, Snipas SJ, Ellerby LM, Bredesen D,
Freeze H, Abrahamson M, Bromme D, Krajewski S, Reed JC, Yin XM, Turk V, Salvesen GS (2001) Lysosomal protease pathways to apoptosis. Cleavage of bid, not pro-caspases, is the most likely
route. _J Biol Chem_ 276: 3149–3157 Article CAS PubMed Google Scholar * Turk B, Stoka V, Rozman-Pungercar J, Cirman T, Droga-Mazovec G, Oreic K, Turk V (2002) Apoptotic pathways:
involvement of lysosomal proteases. _Biol Chem_ 383: 1035–1044 CAS PubMed Google Scholar * Turk B, Turk D, Turk V (2000) Lysosomal cysteine proteases: more than scavengers. _Biochim
Biophys Acta_ 1477: 98–111 Article CAS PubMed Google Scholar * Varghese J, Radhika G, Sarin A (2001) The role of calpain in caspase activation during etoposide induced apoptosis in T
cells. _Eur J Immunol_ 31: 2035–2041 Article CAS PubMed Google Scholar * Yan S, Sameni M, Sloane BF (1998) Cathepsin B and human tumor progression. _Biol Chem_ 379: 113–123 CAS PubMed
Google Scholar Download references ACKNOWLEDGEMENTS A.G. was supported by the SFB 469. This work was supported by Bavarian Government, Roche Diagnostics, and Xantos Biomedicine AG. AUTHOR
INFORMATION AUTHORS AND AFFILIATIONS * Max-Planck-Institute for Biochemistry, Am Klopferspitz 18a, Martinsried, 82152, Germany A Gewies & S Grimm Authors * A Gewies View author
publications You can also search for this author inPubMed Google Scholar * S Grimm View author publications You can also search for this author inPubMed Google Scholar CORRESPONDING AUTHOR
Correspondence to S Grimm. RIGHTS AND PERMISSIONS From twelve months after its original publication, this work is licensed under the Creative Commons Attribution-NonCommercial-Share Alike
3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/ Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Gewies, A., Grimm,
S. Cathepsin-B and cathepsin-L expression levels do not correlate with sensitivity of tumour cells to TNF-_α_-mediated apoptosis. _Br J Cancer_ 89, 1574–1580 (2003).
https://doi.org/10.1038/sj.bjc.6601297 Download citation * Received: 07 May 2003 * Revised: 30 July 2003 * Accepted: 05 August 2003 * Published: 14 October 2003 * Issue Date: 20 October 2003
* DOI: https://doi.org/10.1038/sj.bjc.6601297 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a shareable link is
not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative KEYWORDS * cell death * cathepsin * overexpression * lysosomes