Cycles of transient high-dose cyclophosphamide administration and intratumoral oncolytic adenovirus vector injection for long-term tumor suppression in syrian hamsters
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT Immune responses against oncolytic adenovirus (Ad) vectors are thought to limit vector anti-tumor efficacy. With Syrian hamsters, which are immunocompetent and whose tumors and
normal tissues are permissive for replication of Ad5-based oncolytic Ad vectors, treating with high-dose cyclophosphamide (CP) to suppress the immune system and exert chemotherapeutic
effects enhances Ad vector anti-tumor efficacy. However, long-term CP treatment and immunosuppression can lead to anemia and vector spread to normal tissues. Here, we employed three cycles
of transient high-dose CP administration plus intratumoral injection of the oncolytic Ad vector VRX-007 followed by withdrawal of CP. Each cycle lasted 4–6 weeks. This protocol allowed the
hamsters to remain healthy so the study could be continued for ∼100 days. The tumors were very well suppressed throughout the study. With immunocompetent hamsters, the vector retarded tumor
growth initially, but after 3–4 weeks the tumors resumed rapid growth and further injections of vector were ineffective. Preimmunization of the hamsters with Ad5 prevented vector spillover
from the tumor to the liver yet still allowed for effective long-term anti-tumor efficacy. Our results suggest that a clinical protocol might be developed with cycles of transient
chemotherapy plus intratumoral vector injection to achieve significant anti-tumor efficacy while minimizing the side effects of cytostatic treatment. SIMILAR CONTENT BEING VIEWED BY OTHERS
BETA PRIME: A FIRST-IN-MAN PHASE 1 STUDY OF ADAPT-001, AN ARMED ONCOLYTIC ADENOVIRUS FOR SOLID TUMORS Article Open access 25 December 2023 INTRATUMORAL IMMUNOSTIMULATORY ADCD40L GENE THERAPY
IN PATIENTS WITH ADVANCED SOLID TUMORS Article 14 December 2020 THE SAFETY AND EFFICACY OF SYSTEMIC DELIVERY OF A NEW LIVER-DE-TARGETED TGFΒ SIGNALING INHIBITING ADENOVIRUS IN AN
IMMUNOCOMPETENT TRIPLE NEGATIVE MOUSE MAMMARY TUMOR MODEL Article Open access 24 January 2024 INTRODUCTION Oncolytic adenovirus (Ad) vectors are emerging as a promising form of cancer
therapy (reviewed in refs.1, 2, 3, 4, 5). To be effective, oncolytic viruses need to infect and lyse cancer cells and, in theory, spread efficiently throughout the tumor. Two major barriers
for efficient spread of the vector inside the tumor are hypothesized to include the tumor architecture (physical barrier)6, 7 and the immune system (which clears the vector from the tumor
site).8, 9 We and others have employed the Syrian hamster as a model to study the anti-tumor efficacy, biodistribution and toxicity of oncolytic Ad Serotype 5 (Ad5)-based vectors (reviewed
in refs.10, 11). The virtues of this model are that numerous Syrian hamster tumor cell lines exist that are permissive for Ad5 and that form tumors in hamsters, the tissues of the hamsters
are permissive for Ad5 replication, and the hamsters are immunocompetent. Our long-term goals are to use this model to establish general principles and to develop protocols by which
oncolytic Ad vectors can be used to treat cancer in humans. We9, 12, 13 and others8, 14, 15, 16, 17, 18 have shown that cyclophosphamide (CP) treatment of animals bearing tumors
significantly increases vector anti-tumor efficacy following intratumoral injection of the vector as compared with the untreated counterpart. We used high-dose CP in our studies; the
increased efficacy observed might be due to the immunosuppressive effect of CP that facilitates prolonged persistence and efficient spread of the vector inside the tumor.8, 9, 14 Further,
noting that CP is a chemotherapeutic alkylating agent, we have reported that high-dose CP acts additively with the Ad vector to control tumor growth.12 In our studies, long-term treatment
with CP leads to various side effects such as anemia and increased vector toxicity due to systemic vector spread and replication in normal tissues.9 To minimize the drawbacks of long-term
treatment with CP and still achieve maximum anti-tumor efficacy, in the current study we have employed repeated cycles of transient treatment with CP and vector administration. Subcutaneous
tumors were developed in Syrian hamsters using a hamster renal cancer cell line (HaK), then the hamsters were administered high-dose CP and the tumors were injected with the Ad5-based
oncolytic vector VRX-007. Red blood cell (RBC) and white blood cell (WBC) levels were monitored, and the CP treatment was discontinued temporarily until the RBC recovered. Tumor growth was
measured throughout the study. This protocol mitigated the adverse side effects of CP (for example, anemia) and allowed the study to proceed for ∼100 days. VRX-007 is being evaluated in a
phase I dose escalation clinical trial: intratumoral injection of VRX-007 into advanced solid tumors of any indication (Protocol #510-732). VRX-007 retains all Ad5 genes except those in the
E3 region. To investigate the role of preexisting immunity to Ad5 in vector anti-tumor efficacy and to restrict vector spillover from the tumor to normal tissues in hamsters immunosuppressed
by CP in this long-term study, we immunized the hamsters with Ad5 before CP treatment. Preexisting immunity in the hamster model resembles the situation in many cancer patients inasmuch as
most of the human population in the world has immunity to Ad5.19 We found that tumor growth was effectively suppressed in the preimmunized hamsters and that preimmunization successfully
prevented vector spillover and infection of the liver. MATERIALS AND METHODS ANIMALS Syrian (Golden) hamsters (_Mesocricetus auratus_; 4–5 weeks old) were obtained from Harlan Sprague Dawley
(Indianapolis, IN). Saint Louis University Institutional Animal Care and Use Committee approved the studies. The studies were conducted in accordance with institutional and federal
regulations. VIRUSES AND VECTORS The oncolytic Ad vector VRX-007 is based on Ad5. VRX-007 lacks most of the E3 genes and overexpresses the E3-11.6K Adenovirus Death Protein.20, 21 All other
genes in VRX-007 are intact; despite this, VRX-007 replicates with about 100-fold greater yield in human A549 human lung cancer cells than in primary bronchial epithelial cells22 and it
replicates poorly in quiescent versus proliferating primary human bronchial epithelial cells.23 Introgen Therapeutics (Houston, TX) kindly provided us with the purified stocks.13, 24 VRX-007
is named INGN 007 in some previous publications. IMMUNIZATION Single intramuscular injection of Ad5 (1 × 1010 plaque forming units (p.f.u.)) was used to generate preexisting immunity in
some groups of hamsters 15 days before injecting HaK cells to form tumors.13 ANTITUMOR THERAPY: CYCLES OF TREATMENT WITH CP, INTRATUMORAL INJECTION WITH VRX-007, AND RECOVERY FROM CP
Subcutaneous tumors were established in the hind flank of hamsters by injecting 2 × 107 hamster kidney cancer cells (HaK) in 200 μl of serum-free DMEM.13 When the mean tumor volumes reached
∼600–700 μl, the treatment procedures started. Tumors of ∼600–700 μl are considerably larger than tumors typically studied in animal models using oncolytic viral vectors. The experimental
groups are shown in Table 1. Some groups of hamsters were subjected to the cycles of CP treatment, intratumoral VRX-007 injection, then withdrawal from CP (see Figures 2 and 3). These
hamsters were treated with CP (Sigma-Aldrich, St Louis, MO), and were housed in sterile caging and fed irradiated chow and antibiotic (Baytril Bayer HealthCare (Shawnee Mission, KS)) treated
water.9, 13, 24 The initial dose of CP was 140 mg kg−1 body weight and the subsequent doses were 100 mg kg−1 twice weekly. About 1 week following the first injection of CP and coinciding
with the third CP injection, when the hamsters were severely immunosuppressed, the tumors were administered 5 daily injections of VRX-007, 1 × 1010 p.f.u. per injection. The levels of RBC
and WBC including lymphocytes were monitored by periodic analysis of complete blood counts throughout the length of the study. When the hamsters started to show signs of anemia (less than
normal RBC), the CP was withdrawn and the hamsters were allowed to recover to normal RBC levels. This withdrawal period completed the cycle. Three cycles of CP treatment–VRX-007 intratumoral
injection–withdrawal from CP were completed. The CP-VRX-007 treatment immunosuppression/chemotherapy periods were ∼18–24 days and the recovery periods were ∼14–22 days. Tumor volumes were
measured using digital calipers twice every week. HISTOPATHOLOGY The tumors from each animal were fixed in 10% neutral-buffered formalin. After fixation, the tumors were trimmed, embedded in
paraffin, sectioned (5 μm), and stained with hematoxylin-eosin. A Nikon Optiphot microscope (Nikon, Melville, NY) equipped with a Nikon DXM1200 digital camera and ACT-1 software (Nikon) was
used to capture the images. To obtain a comprehensive view of the tumor, several images were taken from one end of the tumor section to the other end traversing through the center of the
tumor. The images were then stitched together using Adobe Photoshop and Illustrator softwares. NEUTRALIZATION ASSAY Neutralization assay was performed as described previously.13, 24 The
neutralizing antibodies (NAb) titers were determined by the highest dilution of serum/tissue extract that resulted in at least 50% inhibition of cytopathic effect by 100 p.f.u. of VRX-007
assayed on A549 cells. VIRUS QUANTIFICATION IN TISSUES The whole tumor and part of the right lateral lobe of the liver were collected at the time of killing. All the tissues were frozen
immediately in liquid nitrogen and stored at −80 °C. The tissues were weighed and processed as described elsewhere.13, 25 The infectious virus titers were determined by tissue culture
infectious dose 50% (TCID50) assay on A549 cells.13, 25 STATISTICAL ANALYSIS The overall treatment effect was determined by the Kruskal–Wallis test and the Mann–Whitney _U_ test was used for
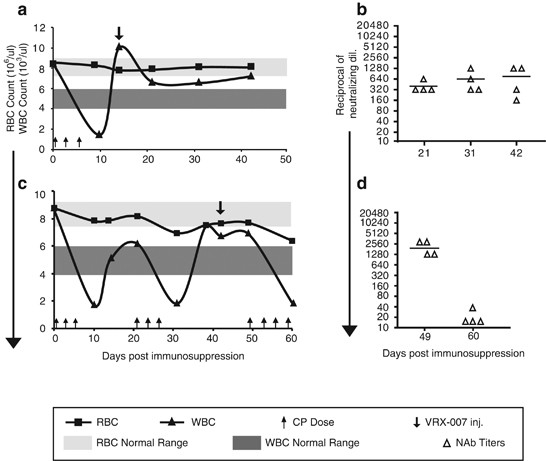
pairwise comparisons. _P_⩽0.05 was considered to be significant. RESULTS KINETICS OF RBC, WBC AND LYMPHOCYTE DECLINE AFTER TREATMENT WITH CP, THE RECOVERY KINETICS AFTER WITHDRAWAL FROM CP,
AND THE GENERATION OF NEUTRALIZING ANTIBODIES AGAINST VRX-007 Before beginning the tumor growth control study, namely cycles of CP and VRX-007 administration, we conducted an experiment to
define the kinetics of RBC, WBC and lymphocyte decline after treatment with CP, the recovery kinetics of these cells after the hamsters were taken off CP, and the generation of NAb against
VRX-007. One group of hamsters was subjected to one round of CP treatment (Figure 1a) and the second group received a second and third round of CP (Figure 1c). VRX-007 was injected
intramuscularly after the first (Figure 1a) or the second (Figure 1c) round of CP and the response of NAb at various days was determined (Figures 1b and d). The WBC (Figure 1a) and
lymphocyte (not shown) counts declined very rapidly by day 10 following the first CP treatment, and the hamsters became severely immunosuppressed. The RBC counts remained within normal range
at 10 days (Figure 1a). The hamsters from the first group were allowed to recover partially from immunosuppression, then VRX-007 was injected at day 14 (that is, 14 days after the first
injection of CP) (Figure 1a). Serum was collected at days 21, 31 and 42, and NAb titers were determined. As shown in Figure 1b, there was a moderate NAb response (∼1:400) at day 21 that
increased to 1:760 at day 42. Thus, the naïve lymphocytes that developed following withdrawal from CP were capable of mounting an NAb response against the vector after only 7 days following
withdrawal from CP. The second group of hamsters was subjected to three rounds of CP (Figure 1c). VRX-007 was injected at day 42, during the lymphocyte recovery period following the second
round of CP (Figure 1c). At 7 days following VRX-007 injection, the hamsters were treated a third time with CP, and the lymphocytes declined again. A good NAb response was observed at day 49
(∼1:1920) that probably was generated by lymphocytes between day 41 and 49 when CP was administered again. Little or no NAb was detected by day 60 (Figure 1d). Importantly, the RBC levels
for the most part recovered when the CP was withdrawn (Figures 1a and c). As such, the hamsters did not become anemic. In summary, the levels and function of RBC and lymphocytes can be
controlled by using rounds of CP treatment. Lymphocytes formed following CP withdrawal are able to generate NAb within 7 days of VRX-007 intramuscular injection. DESIGN OF THE LONG-TERM
TUMOR GROWTH CONTROL STUDY We next conducted the tumor growth control study being guided in part by the findings from Figure 1. As part of the study, we wished to evaluate the anti-tumor
efficacy of intratumoral VRX-007 in immunocompetent hamsters, the anti-tumor efficacy of cycles of treatment with CP and VRX-007, whether the cycling of CP mitigated anemia, whether cycling
controlled the levels of WBC, and the role of preexisting NAb to the vector. Six groups of hamsters bearing subcutaneous HaK tumors were established. The nature of these groups is depicted
in Table 1. These groups and the results obtained will be described in the following sections. The procedure used for the multiple cycles of transient treatment with CP followed by VRX-007
intratumoral administration is shown in Figures 2 and 3 and is described in the legend of Figure 3. The study lasted ∼100 days following the first treatment with CP. During the entire study
the RBC and WBC levels were monitored closely (Figure 2), and were used to determine the duration of CP treatment and the recovery period. The tumor growth measurements are in Figure 3. For
ease of presentation and discussion, the cumulative tumor growth is shown after the first (Figure 3a), second (Figure 3b) and third (Figure 3c) cycles of CP and vector administration. Tumors
from an arm of the experiment were harvested at days 43, 74 and 90 and the NAb titers and the amount of VRX-007 in the tumors were determined (Figure 4). Histopathology of typical
hematoxylin-eosin-stained tumors at day 46 is in Figure 5. VRX-007 IS EFFECTIVE FOR A PERIOD IN IMMUNOCOMPETENT HAMSTERS, BUT THEN TUMOR REGROWTH OCCURS AND A SECOND SERIES OF VECTOR
INJECTION IS NOT EFFECTIVE As shown in Figure 3a, growth of tumors was rapid in preimmunized hamsters that were immunocompetent and treated with buffer alone (Imm+Buffer) or hamsters that
were immunosuppressed with CP and treated with buffer (Imm+CP+Buffer). The CP treatment retarded tumor growth slightly, but it was not statistically significant (_P_=0.347) (Figure 3a). The
buffer-injected groups were terminated at 28 days because of their large tumor burden. Injection of VRX-007 into tumors of immunocompetent hamsters (NonImm+007, Imm+007) retarded tumor
growth for 3–4 weeks compared with the buffer control (_P_=0.005) (Figure 3a). However, the vector-injected tumors then began to grow rapidly (Figure 3b). Notably, a second round of
intratumoral vector injection (5 consecutive daily injections of 1 × 1010 p.f.u. per injection, at days 29–33) did not affect the increased tumor growth rate (Figure 3b). These hamsters were
terminated at day 46. It is important to understand why VRX-007 ceased to be effective in these immunocompetent hamsters. One reason could be vector elimination by NAb. Indeed, anti-Ad5 NAb
was very high in the tumors at day 43 (∼1:40 960 in the Imm+007 group, ∼1:10 240 in the Non-Imm+007 group) (Figure 4a). Not surprisingly from these high NAb levels, vector could not be
detected in the tumors of these groups (Figure 4b). Those tumors from animals that were not treated with CP contained mostly viable cells (Figure 5, top two panels), indicating that the
tumors were growing rapidly. CP PLUS VRX-007 CAN BE ADMINISTERED IN MULTIPLE CYCLES AND ARE STRONGLY SUPPRESSIVE OF TUMOR GROWTH The following discussion pertains to the groups treated with
CP plus vector. First, considering the preimmunized group (Imm+CP+007), at 28 days, the tumors grew faster than for the NonImm+CP+007 group (_P_=0.015) (Figure 3a). This lesser vector
efficacy in the Imm+CP+007 group was likely due to the presence of NAbs inside the tumor at the time of vector administration as described in our previous studies.13 After the second cycle
of CP treatment–vector injection–recovery, tumors in both CP groups (Imm+CP+007 and Non-Imm+CP+007) were controlled much better than the tumors in the non-CP groups injected with VRX-007
(_P_=0.000) (Figure 3b). Clearly, the combination of high-dose CP treatment plus vector is much superior to either CP alone or vector alone. The question arises whether CP is enhancing
vector replication and persistence though CP’s immunosuppressive activity and/or whether CP is acting as a chemotherapeutic agent. It is noteworthy that after the second cycle of CP–vector
injection–recovery the tumors in the Imm+CP+007 group were controlled equally as well as in the NonImm+CP+007 group (_P_=0.581) (Figure 3b). As shown in Figure 4a, at day 43, NAb was found
in the tumors of the Imm+CP+007 group (∼1:320) and the NonImm+CP+007 group (∼1:40). No vector was detected in the tumors of these groups. It should be noted that the data in Figures 4a and b
are from tumors collected 14 days after the last vector injection, and also that the tumors at this point consisted mostly of necrotic areas and with limited patches of viable cells (see
Figures 5c and d). No doubt there was vector in the tumors at the time of injection; the lack of vector in the tumors at day 43 is probably due to a combination of vector neutralization by
NAb and a paucity of viable cells in the tumors from the CP-treated hamsters in which the vector can replicate. In any case, the suppression of tumor growth in the absence of vector suggests
that CP is having a chemotherapeutic effect over the period days 28–46. Starting at about day 65, tumors in both CP+vector groups started to grow again (Figure 3c). This could be because
the tumors were devoid of vector (that is, there was no vector at day 43, see Figure 4b). Also, the hamsters had been off CP for 18 days, so any chemotherapeutic effect due to CP might be
waning. A third cycle of CP treatment was administered at days 63–88, and vector was injected for a third round at days 69–73 (Figure 3c). This CP+vector treatment seemed to stop the
regrowth of tumors and cause a decrease in tumor size (Figure 3c). Measurement of tumors continued until day 90 following the first injection of vector. Remarkably, the tumors in both
CP-treated groups were controlled very well and equally well (_P_=0.190) with a tumor volume almost equal to the starting volume. Again, the question arises, what is responsible for
retarding the tumor growth: the vector, CP or both? At day 74, 1 day after the last injection of vector in the third cycle of vector injection, high levels of vector (108 TCID50 units) were
detected in the tumors of the NonImm+CP+007 group, and much lower levels (∼103 TCID50 units) in the Imm+CP+007 group (Figure 4d). At day 90, 18 days after the last injection of vector,
vector (∼105 TCID50 units) continued to be isolated from tumors in the NonImm+CP+007 group, but not from the Imm+CP+007 group (Figure 4f). The tumors of the Imm+CP+007 group seemed to have
more NAb than the NonImm+CP+007 group at day 74 (Figure 4c) and day 90 (Figure 4e), and that could explain why the Imm+CP+007 tumors had less vector. In any case, considering that the
NonImm+CP+007 group had the most vector in the tumor yet had similar growth suppression as the Imm+CP+007 group, it seems possible that the tumor growth suppression may be due to CP
chemotherapy. However, the vector injected at days 69–73 likely contributed as well to suppression of tumor growth. Even though the tumors in the hamsters were very well controlled, some
animals died during the late phases of the study. Upon necropsy, it was found that the subcutaneous tumor had metastasized to several tissues such as lungs and kidney that might be the
reason for the deaths (Supplementary Figure 1). Therefore, we terminated the experiment at day 90 post first vector injection. Figure 5 shows the histopathology analysis of representative
tumors at day 46 of the experiment. The tumors from the non-CP-treated groups appeared to have viable cells (purple stained cells) throughout much of the tumor (Figures 5a and b), which
explains why these tumors were growing rapidly (Figure 3b). As discussed, the high NAb in these tumors likely eliminated the vector. The tumors from the CP+VRX-007 groups had very few viable
cells and large necrotic areas after the second cycle (Figures 5c and d) and also after the third cycle (data not shown). The paucity of viable cells in these latter tumors may explain why
these tumors had little vector remaining (Figure 4b) from the last injection of vector 10 days previously, and why these tumors were growing slowly, if at all. Notably, the tumor mass that
we were measuring using digital calipers (Figure 3) in the hamsters that underwent cycling comprised mainly dead tissue. SERUM NEUTRALIZING ANTIBODIES PREVENT VRX-007 SPILLOVER FROM THE
TUMOR TO THE LIVER We investigated whether serum NAb could minimize vector spillover from tumors. As shown in Figure 4g for the Imm+CP+007 group, preimmunization effectively prevented vector
spillover from the tumor to the liver at all time points (days 43, 74 and 90). The naïve hamsters (NonImm+CP+007), on the other hand, had vector in the liver: ∼106 TCID50 per gram of liver
at day 43 and ∼3 × 104 TCID50 per gram of liver at day 74 (Figure 4g). No vector was detected in the liver at day 90 (kill). DISCUSSION A notable aspect of our results is that, in
immunocompetent hamsters, VRX-007 injected intratumorally is able to retard tumor growth for 3–4 weeks, but then tumors start to grow rapidly. A second round of vector injection has no
effect on tumor growth, probably because there are high levels of NAb in the tumor. These results are important considering that clinical protocols usually recommend multiple rounds of
vector injection into patients that are for the most part immunocompetent.26, 27 Another notable aspect of our results is that the combination of intratumoral injection with VRX-007 plus
high-dose CP treatment is much more effective at suppressing tumor growth than either VRX-007 alone or CP alone. With our cycling protocol, we were able to achieve very effective long-term
control of tumor growth due to aggressive virotherapy (high dose of VRX-007) and chemotherapy (high dose of CP), while mitigating, at least to some degree, the toxic effects of high-dose CP.
This tumor growth suppression was achieved even though the starting tumors were large (600–700 μl) and vascularized. Another aspect of our results is that preimmunization of the hamsters
with Ad5 did not affect the long-term anti-tumor efficacy of the VRX-007+CP combination. Importantly, preimmunization was able to prevent spillover of VRX-007 from the tumors to the liver
over the course of 90 days following first intratumoral injection of vector in the hamsters that were treated with CP. CP has been employed by a number of laboratories to increase the
anti-tumor efficacy and/or to decrease the immune response to oncolytic vectors including those based on Ad,9, 12, 13, 15, 24, 28, 29 herpes simplex virus type 1,8, 16 reovirus,14 measles
virus,17, 30 vesicular stomatitis virus30 and vaccinia virus.18 A variety of CP doses and dosing regimes have been used, usually in mouse models. Peng _et al_30 concluded that 120 mg kg−1 of
CP given daily by intraperitoneal injection for 4 days was a good regime to control the humoral response to intravenously administered oncolytic measles virus or vesicular stomatitis virus,
but they did not study suppression of tumor growth. They noted that this regime was similar to CP dosing regimes used in humans for chemotherapy of cancer. With oncolytic Ads in the Syrian
hamster tumor model, low-dose CP, for example, 20 mg kg−1 intraperitoneally twice per week31 or 300 or 500 mg kg−1 given once intraperitoneally,28 has been used as a means to specifically
delete regulatory T cells. Low-dose CP, for example, metronomic dosing of 50 mg kg−1, has been used in cancer patients in combination with oncolytic Ad vectors.28, 31 Another laboratory15
working with an oncolytic Ad vector in the Syrian hamster model reported the successful use of intermediate-dose CP (37 mg kg−1, twice weekly) as well as the high-dose regime used in our
current study. As discussed above, high-dose CP administered to animal models is toxic. However, our results reported here suggest that cycles of transient chemotherapy (and
immunosuppression) using CP and vector intratumoral administration is a promising approach, one that has not been attempted previously in an immunocompetent animal model that is permissive
for replication of oncolytic Ad vectors. The procedure allowed us to control the tumor growth and to keep the hamsters healthier for a much longer time than in previous studies. We speculate
from our results that a clinical protocol might be developed with cycles of transient chemotherapy coupled with multiple rounds of intratumoral vector injection to achieve significant
antitumor efficacy while minimizing the side effects of chemotherapy and vector. Chemotherapy regimens involve repeated cycles of dosing of the chemotherapy drug followed by recovery similar
to the protocol employed by us in the current study. We used high-dose CP, but an intermediate dose such as 37 mg kg−1, twice weekly, might be sufficient.15 Also, other chemotherapeutic
agents could work as well as many of these cause transient immunosuppression. To prevent vector spread, patients could either be immunized with a replication-defective Ad before the
treatment, screened for preexisting anti-Ad NAb, or patients could be passively immunized with anti-Ad NAb.24 Chemotherapy has shown to increase the vector efficacy when used together.32
REFERENCES * Wold WSM, Ison MG . Adenoviruses. In: Knipe DM, Howley PM editors. _Fields Virology_. 6th edition. Lippincott Williams & Wilkins Philadelphia, PA, 2013: 1732–1767. Google
Scholar * Eager RM, Nemunaitis J . Clinical development directions in oncolytic viral therapy. _Cancer Gene Ther_ 2011; 18: 305–317. Article CAS PubMed Google Scholar * Toth K, Dhar D,
Wold WSM . Oncolytic (replication-competent) adenoviruses as anticancer agents. _Expert Opin Biol Ther_ 2010; 10: 353–368. Article CAS PubMed Google Scholar * Pesonen S, Kangasniemi L,
Hemminki A . Oncolytic adenoviruses for the treatment of human cancer: focus on translational and clinical data. _Mol Pharm_ 2011; 8: 12–28. Article CAS PubMed Google Scholar * Shi J,
Zheng D . An update on gene therapy in China. _Curr Opin Mol Ther_ 2009; 11: 547–553. CAS PubMed Google Scholar * Ganesh S, Gonzalez EM, Idamakanti N, Abramova M, Vanroey M, Robinson M
_et al_. Relaxin-expressing, fiber chimeric oncolytic adenovirus prolongs survival of tumor-bearing mice. _Cancer Res_ 2007; 67: 4399–4407 [Erratum appears in Cancer Res. 2007;67(12):5998].
Article CAS PubMed Google Scholar * McKee TD, Grandi P, Mok W, Alexandrakis G, Insin N, Zimmer JP _et al_. Degradation of fibrillar collagen in a human melanoma xenograft improves the
efficacy of an oncolytic herpes simplex virus vector. _Cancer Res_ 2006; 66: 2509–2513. Article CAS PubMed Google Scholar * Fulci G, Breymann L, Gianni D, Kurozomi K, Rhee SS, Yu J _et
al_. Cyclophosphamide enhances glioma virotherapy by inhibiting innate immune responses. _Proc Natl Acad Sci USA_ 2006; 103: 12873–12878. Article CAS PubMed PubMed Central Google Scholar
* Thomas MA, Spencer JF, Toth K, Sagartz JE, Phillips N, Wold WSM . Immunosuppression enhances oncolytic adenovirus replication and anti tumor efficacy in the Syrian hamster model. _Mol
Ther_ 2008; 16: 1665–1673. Article CAS PubMed Google Scholar * Wold WS, Toth K . Chapter three—Syrian hamster as an animal model to study oncolytic adenoviruses and to evaluate the
efficacy of antiviral compounds. _Adv Cancer Res_ 2012; 115: 69–92. Article CAS PubMed Google Scholar * Dhar D, Toth K, Wold WSM . Syrian hamster tumor model to study oncolytic Ad5 based
vectors. In: Kirn D, Liu TC, Thorne S editors. _Oncolytic Viruses: Methods and Protocols_. Humana Press New York, 2012; 797: 53–63. Chapter Google Scholar * Young BA, Spencer JF, Ying B,
Tollefson AE, Toth K, Wold WSM . The role of cyclophosphamide in enhancing antitumor efficacy of an adenovirus oncolytic vector in subcutaneous Syrian hamster tumors. _Cancer Gene Ther_
2013; 20: 521–530. Article CAS PubMed PubMed Central Google Scholar * Dhar D, Spencer JF, Toth K, Wold WSM . Effect of preexisting immunity on oncolytic adenovirus vector INGN 007
antitumor efficacy in immunocompetent and immunosuppressed Syrian hamsters. _J Virol_ 2009; 83: 2130–2139. Article CAS PubMed Google Scholar * Qiao J, Wang H, Kottke T, White C, Twigger
K, Diaz RM _et al_. Cyclophosphamide facilitates antitumor efficacy against subcutaneous tumors following intravenous delivery of reovirus. _Clin Cancer Res_ 2008; 14: 259–269. Article CAS
PubMed PubMed Central Google Scholar * Hasegawa N, Abei M, Yokoyama KK, Fukuda K, Seo E, Kawashima R _et al_. Cyclophosphamide enhances antitumor efficacy of oncolytic adenovirus
expressing uracil phosphoribosyltransferase (UPRT) in immunocompetent Syrian hamsters. _Int J Cancer_ 2013; 133: 1479–1488. Article CAS PubMed Google Scholar * Ikeda K, Ichikawa T,
Wakimoto H, Silver JS, Deisboeck TS, Finkelstein D _et al_. Oncolytic virus therapy of multiple tumors in the brain requires suppression of innate and elicited antiviral responses. _Nat Med_
1999; 5: 881–887. Article CAS PubMed Google Scholar * Ungerechts G, Springfeld C, Frenzke ME, Lampe J, Parker WB, Sorscher EJ _et al_. An immunocompetent murine model for oncolysis with
an armed and targeted measles virus. _Mol Ther_ 2007; 15: 1991–1997. Article CAS PubMed Google Scholar * Lun XQ, Jang JH, Tang N, Deng H, Head R, Bell JC _et al_. Efficacy of
systemically administered oncolytic vaccinia virotherapy for malignant gliomas is enhanced by combination therapy with rapamycin or cyclophosphamide. _Clin Cancer Res_ 2009; 15: 2777–2788.
Article CAS PubMed Google Scholar * Chirmule N, Propert K, Magosin S, Qian Y, Qian R, Wilson J . Immune responses to adenovirus and adeno-associated virus in humans. _Gene Ther_ 1999; 6:
1574–1583. Article CAS PubMed Google Scholar * Doronin K, Toth K, Kuppuswamy M, Krajcsi P, Tollefson AE, Wold WSM . Overexpression of the ADP (E3-11.6K) protein increases cell lysis and
spread of adenovirus. _Virology_ 2003; 305: 378–387. Article CAS PubMed Google Scholar * Tollefson AE, Scaria A, Hermiston TW, Ryerse JS, Wold LJ, Wold WSM . The adenovirus death
protein (E3-11.6K) is required at very late stages of infection for efficient cell lysis and release of adenovirus from infected cells. _J Virol_ 1996; 70: 2296–2306. CAS PubMed PubMed
Central Google Scholar * Kuppuswamy M, Spencer JF, Doronin K, Tollefson AE, Wold WS, Toth K . Oncolytic adenovirus that overproduces ADP and replicates selectively in tumors due to hTERT
promoter-regulated E4 gene expression. _Gene Ther_ 2005; 12: 1608–1617. Article CAS PubMed Google Scholar * Toth K, Kuppuswamy M, Shashkova EV, Spencer JF, Wold WSM . A fully
replication-competent adenovirus vector with enhanced oncolytic properties. _Cancer Gene Ther_ 2010; 17: 761–770. Article CAS PubMed PubMed Central Google Scholar * Dhar D, Spencer JF,
Toth K, Wold WSM . Pre-existing immunity and passive immunity to Adenovirus 5 prevents toxicity caused by an oncolytic adenovirus vector in the Syrian Hamster Model. _Mol Ther_ 2009; 17:
1724–1732. Article CAS PubMed PubMed Central Google Scholar * Thomas MA, Spencer JF, Wold WSM . Use of the Syrian hamster as an animal model for oncolytic adenovirus vectors. _Methods
Mol Med_ 2007; 130: 169–183. CAS PubMed Google Scholar * Chiocca EA, Abbed KM, Tatter S, Louis DN, Hochberg FH, Barker F _et al_. A phase I open-label, dose-escalation,
multi-institutional trial of injection with an E1B-attenuated adenovirus, ONYX-015, into the peritumoral region of recurrent malignant gliomas, in the adjuvant setting. _Mol Ther_ 2004; 10:
958–966. Article CAS PubMed Google Scholar * Galanis E, Okuno SH, Nascimento AG, Lewis BD, Lee RA, Oliveira AM _et al_. PhaseI-II trial of ONYX-015 in combination with MAP chemotherapy
in patients with advanced sarcomas. _Gene Ther_ 2005; 12: 437–445. Article CAS PubMed Google Scholar * Cerullo V, Diaconu I, Kangasniemi L, Rajecki M, Escutenaire S, Koski A _et al_.
Immunological effects of low-dose cyclophosphamide in cancer patients treated with oncolytic Adenovirus. _Mol Ther_ 2011; 19: 1737–1746. Article CAS PubMed PubMed Central Google Scholar
* Lamfers ML, Fulci G, Gianni D, Tang Y, Kurozumi K, Kaur B _et al_. Cyclophosphamide increases transgene expression mediated by an oncolytic adenovirus in glioma-bearing mice monitored by
bioluminescence imaging. _Mol Ther_ 2006; 14: 779–788. Article CAS PubMed Google Scholar * Peng KW, Myers R, Greenslade A, Mader E, Greiner S, Federspiel MJ _et al_. Using clinically
approved cyclophosphamide regimens to control the humoral immune response to oncolytic viruses. _Gene Ther_ 2013; 20: 255–261. Article CAS PubMed Google Scholar * Koski A, Kangasniemi L,
Escutenaire S, Pesonen S, Cerullo V, Diaconu I _et al_. Treatment of cancer patients with a serotype 5/3 chimeric oncolytic adenovirus expressing GMCSF. _Mol Ther_ 2010; 18: 1874–1884.
Article CAS PubMed PubMed Central Google Scholar * Khuri FR, Nemunaitis J, Ganly I, Arseneau J, Tannock IF, Romel L _et al_. A controlled trial of intratumoral ONYX-015, a selectively
replicating adenovirus, in combination with cisplatin and 5-fluorouracil in patients with recurrent head and neck cancer. _Nat Med_ 2000; 6: 879–885. Article CAS PubMed Google Scholar
Download references ACKNOWLEDGEMENTS We thank Jacqueline F Spencer for her help in bleeding the animals. This research was supported by grant CA118022 to WSMW from the National Institutes of
Health. AUTHOR INFORMATION Author notes * D Dhar Present address: 2Current address: Dr Dhar, Laboratory of Gene Regulation and Signal Transduction, Department of Pharmacology, University of
California, San Diego, School of Medicine, 9500 Gilman Drive, Leichtag Building, MC 0723, La Jolla, CA 92093, USA, AUTHORS AND AFFILIATIONS * Department of Molecular Microbiology and
Immunology, Saint Louis University School of Medicine, St Louis, MO, USA D Dhar, K Toth & W S M Wold Authors * D Dhar View author publications You can also search for this author
inPubMed Google Scholar * K Toth View author publications You can also search for this author inPubMed Google Scholar * W S M Wold View author publications You can also search for this
author inPubMed Google Scholar CORRESPONDING AUTHOR Correspondence to W S M Wold. ETHICS DECLARATIONS COMPETING INTERESTS The VRX-007 vector, which is owned by VirRx, is being evaluated in a
Phase I clinical trial. WSMW and KT own stock in VirRx. ADDITIONAL INFORMATION Supplementary Information accompanies the paper on Cancer Gene Therapy website SUPPLEMENTARY INFORMATION
SUPPLEMENTARY FIGURE S1 (JPG 217 KB) SUPPLEMENTARY FIGURE LEGEND (DOC 29 KB) RIGHTS AND PERMISSIONS Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Dhar, D., Toth, K. &
Wold, W. Cycles of transient high-dose cyclophosphamide administration and intratumoral oncolytic adenovirus vector injection for long-term tumor suppression in Syrian hamsters. _Cancer Gene
Ther_ 21, 171–178 (2014). https://doi.org/10.1038/cgt.2014.13 Download citation * Received: 31 October 2013 * Revised: 06 March 2014 * Accepted: 10 March 2014 * Published: 11 April 2014 *
Issue Date: April 2014 * DOI: https://doi.org/10.1038/cgt.2014.13 SHARE THIS ARTICLE Anyone you share the following link with will be able to read this content: Get shareable link Sorry, a
shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing initiative