Gut microbiota develop towards an adult profile in a sex-specific manner during puberty
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT Accumulating evidence indicates that gut microbiota may regulate sex-hormone levels in the host, with effects on reproductive health. Very little is known about the development of
intestinal microbiota during puberty in humans. To assess the connection between pubertal timing and fecal microbiota, and to assess how fecal microbiota develop during puberty in comparison
with adult microbiota, we utilized a Finnish allergy-prevention-trial cohort (Flora). Data collected at 13-year follow-up were compared with adult data from a different Finnish cohort.
Among the 13-year-old participants we collected questionnaire information, growth data from school-health-system records and fecal samples from 148 participants. Reference adult fecal
samples were received from the Health and Early Life Microbiota (HELMi) cohort (n = 840). Fecal microbiota were analyzed using 16S rRNA gene amplicon sequencing; the data were correlated
with pubertal timing and compared with data on adult microbiota. Probiotic intervention in the allergy-prevention-trial cohort was considered as a confounding factor only. The main outcome
was composition of the microbiota in relation to pubertal timing (time to/from peak growth velocity) in both sexes separately, and similarity to adult microbiota. In girls, fecal microbiota
became more adult-like with pubertal progression (p = 0.009). No such development was observed in boys (p = 0.9). Both sexes showed a trend towards increasing relative abundance of
estrogen-metabolizing Clostridia and decreasing Bacteroidia with pubertal development, but this was statistically significant in girls only (p = 0.03). In girls, pubertal timing was
associated positively with exposure to cephalosporins prior to the age of 10. Our data support the hypothesis that gut microbiota, particularly members of _Ruminococcaceae_, may affect
pubertal timing, possibly via regulating host sex-hormone levels. TRIAL REGISTRATION The registration number for the allergy-prevention-trial cohort: ClinicalTrials.gov, NCT00298337,
registered 1 March 2006—Retrospectively registered, https://clinicaltrials.gov/show/NCT00298337. The adult-comparison cohort (HELMi) is NCT03996304. SIMILAR CONTENT BEING VIEWED BY OTHERS
DELAYED GUT MICROBIOTA MATURATION IN THE FIRST YEAR OF LIFE IS A HALLMARK OF PEDIATRIC ALLERGIC DISEASE Article Open access 29 August 2023 DEVELOPMENT OF GUT MICROBIOTA DURING THE FIRST 2
YEARS OF LIFE Article Open access 31 May 2022 THE BREAST MILK AND CHILDHOOD GASTROINTESTINAL MICROBIOTAS AND DISEASE OUTCOMES: A LONGITUDINAL STUDY Article Open access 10 October 2022
INTRODUCTION The onset of puberty is affected by genetic and environmental factors. Currently, variation in several hundred chromosomal loci are known to be associated with variation in the
onset of puberty1, and external and internal cues such as diet2, exercise3, the amount of fat tissue4, intrauterine conditions5, psychosocial stress6, and chronic diseases7 are known to play
a role. There is rapidly growing evidence pointing to the role of gut microbiota in the regulation of growth via hormonal effects on bone growth8, widespread metabolic effects9 and effects
on nutritional status10. Infant gut microbiota predict body mass index (BMI) at preschool age, and more specifically, _Streptococcus_ and _Bacteroides_ spp. are positively and bifidobacteria
negatively connected to later BMI11. Modulation of gut microbiota in order to improve growth is routine practice in animal husbandry12. Gut microbiota may thus influence pubertal
development via metabolic effects. Gut microbiota are important regulators of circulating estrogens. Conjugated estrogen is secreted to bile, and certain gut microbes, particularly
_Ruminococcus_ and _Faecalibacterium_ spp. secrete beta-glucuronidase, which deconjugates estrogen back to an active form13. Through the enterohepatic circulation, the deconjugated estrogens
return to the systemic circulation. Several human studies have shown associations between the composition of gut microbiota and urinary and fecal estrogen levels14,15. In monkeys, a
decrease of estrogen-metabolizing bacteria leads to a decrease in circulating estrogens16. In addition, dietary fiber consumption, which regulates the composition of the gut microbiota,
affects the levels of serum estrogens17,18,19, suggesting an effect of gut microbiota on sex-hormone levels. Sex hormones in connection with the composition of gut microbiota have been
studied most thoroughly in mice, where gonadectomy changes the composition of the microbiota20, indicating an effect of sex hormones. Furthermore, it has been shown that the gut microbiota
of mice influence sex-hormone levels, which in turn affects the hosts’ immune development21,22. In mice, gut microbiota undergo sex-specific changes during puberty that influence hormone
levels22. Furthermore, fecal microbiota transplants (FMTs) from healthy female rats to those with polycystic ovary syndrome (PCOS) have been shown to ameliorate the condition23, indicating
that the gut microbiota have a decisive impact on gonadal functioning in the host. Accumulating evidence thus suggests that gut microbiota may have an important role in determining the
timing of puberty via metabolic and hormonal effects. It is also possible that gut microbiota composition is dependent on hormonal signals. It has been reported that girls with idiopathic
central precocious puberty (ICPP) have different gut microbiota compared with controls, especially as regards _Ruminococcus_ and _Gemmiger_ spp., which were found to be enriched in the ICPP
group. The microbiota of ICPP patients were also more generally diverse and showed features that have been previously associated with the microbiota of obese individuals24. Limited data
exist on human adolescent gut microbiota. In a recent cross-sectional investigation it was discovered that the distinction in gut microbiota between sexes becomes more evident in puberty25.
Differences have been found between adolescent and adult microbiota, and the amounts of bifidobacteria in particular have been found to diminish with age in several studies26,27,28,29, and
age-related associations with Bacteroidetes and Firmicutes have also been reported27,28,29. However, significant individual variation exists in pubertal timing, and previous studies have not
addressed the associations between pubertal stage and gut microbiota. Hence, we here investigated the association between intestinal microbiota and pubertal timing in humans in a
well-characterized and longitudinally monitored cohort. METHODS PARTICIPANTS AND DATA SOURCES The study was implemented among an allergy-prevention-trial cohort including 1018 mother–child
pairs where the child had a high risk of allergy30 (trial registration number NCT 00298337). The subjects randomly received a mixture of four probiotics in capsules: _Lactobacillus
rhamnosus_ GG(ATCC 53103), 5 × 109 colony-forming units (cfu); _L. rhamnosus_ LC705(DSM 7061), 5 × 10^9 cfu; _Bifidobacterium breve_ Bb99(DSM 13692), 2 × 108 cfu; and _Propionibacterium
freudenreichii_ ssp. shermanii JS(DSM 7076), 2 × 109 cfu, or placebo. Starting 2–4 weeks before the end of pregnancy, mothers took capsules twice daily and children from day 1–2, one daily
capsule and a prebiotic oligosaccharide, or placebo for the first six months of life. The treatment had no effect on growth31. At the age of 13 years, 960 participants were invited for a
follow-up visit. For the visit, 642 participants filled in questionnaires with information on gastrointestinal symptoms and 422 participants provided a fecal sample. Growth data on 306
participants was obtained from school health-service records32. Data on lifetime antibiotic use were obtained from the drug-purchase register of the Finnish National Health Insurance scheme
(Kela) for all participants (Suppl. Figure 1). Written informed consent was obtained from all subjects’ parents and/or legal guardians. The study was approved by the Helsinki University
Hospital Ethics Committee. The ethical statement number is 78/13/03/03/2013. Both growth data and fecal samples were available for 65 boys and 83 girls (total 148). This sub-set was used for
the majority of the analyses. Gastrointestinal symptom data were missing for 4 boys and 2 girls and antibiotic purchase data were missing for 3 girls, so the complete data was available for
61 boys and 78 girls (total 139). Fecal samples from adults of reproductive age were obtained from Finnish Health and Early Life Microbiota (HELMi) cohort33, and used as reference material.
A total of 396 samples from women and 444 samples from men were included. The adult reference cohort consisted of generally healthy parents of newborn infants. DETERMINATION OF THE TIMING
OF PUBERTY FROM GROWTH DATA Reference growth-velocity data on prepubertal growth was obtained from previous work34. In total, 1127 pubertal (over eight years for girls and over 9 years for
boys) measurements were available and the mean number of measurements per person was 7.6. Age at take-off of pubertal growth acceleration was defined as the age when growth velocity exceeded
the age- and sex-specific mean plus 2 SD for the first time, followed by a period of accelerated growth typical for puberty. Age at peak-height velocity (APHV) was defined as the age when a
participant’s growth velocity was highest during a phase of accelerated growth after eight and nine years of age in girls and boys, respectively. Only measurements that were at least six
months apart were used for growth-velocity calculation. For 11 boys and 23 girls, it was only possible to determine APHV according to the aforementioned criteria. In these cases, the age at
take-off was imputed to be six months prior to APHV. To assess the correlation between gut microbiota and the timing of puberty, two new variables were derived. First, the time between
growth take-off and sample collection, and second, the time between APHV and sample collection. FECAL SAMPLE ANALYSIS Fecal samples were collected by the participants at home and frozen
immediately. They were transported to the laboratory frozen and stored at − 80 °C until processing. Bacterial DNA was extracted using a previously described repeated bead-beating method35
with the following modifications for automated DNA purification: ca. 125 mg of fecal material were suspended in 1 ml of sterile ice-cold PBS, and 175 μl of fecal suspension was combined with
235 μl of RBB lysis buffer (500 mM NaCl, 50 mM Tris–HCl (pH 8.0), 50 mM EDTA, 4% SDS) in a bead-beating tube from the Ambion Magmax™ Total Nucleic Acid Isolation Kit (Life Technologies,
Carlsbad, CA, USA). After repeated bead-beating, 200 μl of the supernatant was used for DNA extraction with a KingFisher™ Flex automated purification system (ThermoFisher Scientific,
Waltham, MA, USA) using a MagMAX™ Pathogen High Vol. Duo program. DNA was quantified using a Quanti-iT™ Pico Green dsDNA assay (Invitrogen, San Diego, CA, USA) and 1 ng was used for
V3–V4-region amplicon PCR of the 16S rRNA gene as previously described36. Sequencing was carried out with Illumina HiSeq 2500 equipment in Rapid Run mode. STATISTICAL ANALYSIS AND
VISUALIZATION The sequencing reads were processed using the ProcessReads and TaxonomicTables functions in the R package mare37. The forward reads were quality and chimera filtered using
USEARCH38 v 8.1 and mapped to the Silva39 reference database. After mapping the reads to the reference database, the reads were summarized at different taxonomic levels. Statistical analysis
was conducted in R40, partly using the R package mare, which relies on the R packages vegan41, nlme42, and MASS43. Plots and other figures were made with R40 using the R package ggplot244
and Microsoft Powerpoint for Mac45. Similarity to microbiota in adults was assessed as the correlation between the log-transformed relative abundances of the bacterial taxa at species level
between each participant and an adult of the same sex. The average similarity to adults for each participant was taken as a measure of microbiota maturity. Associations between bacterial
taxa and pubertal timing were analyzed using regression as described below. The number of reads per taxon was used as the response variable and the model was adjusted for probiotic use, time
since the last antibiotic course, BMI and whole-grain intake as a proxy of dietary-fiber intake. The total number of reads per sample was used as an offset. For each taxon, a suitable model
was identified based on standard statistical diagnostics. Initially, a generalized linear model (glm) with negative binomial distribution was fitted. In cases in which the model did not
converge, a glm with Poisson distribution was fitted, and if that did not converge, finally a linear model (lm) using relative abundances instead of raw read counts was fitted
(log-transformed if necessary depending on the data distribution). In cases in which there were patterns in the residuals, a generalized least squares (gls) model was fitted, also modelling
the residual variation. If no model fulfilled standard statistical diagnostic criteria (normality and stochasticity of residuals), no p-value was obtained. Using this iterative approach, a
suitable statistical model was identified for each taxon. RESULTS Comparison of the whole allergy-prevention-trial-cohort at 13-years and the analysed cohort is presented in Suppl Table 1.
The groups were reasonably similar, except for the prevalence of asthma (16.8% in whole cohort vs 7.2% in the analysed cohort). The difference was statistically significant (p = 0.004). The
birth weight in whole cohort (average 3588 g) was higher compared to analyzed cohort (average 3452 g) and the difference was statistically significant (p = 0.002). At 13 years of age, the
boys, as expected, were less mature in terms of pubertal development than the girls (Suppl. Figure 2). Girls were mostly (81%) past their peak-growth velocity, while most boys (80%) had not
yet reached it. Self-reported Tanner staging reflected a similar picture (Suppl. Table 2). Gut microbiota were analyzed in fecal samples by extracting DNA, amplifying the 16S rRNA gene and
sequencing the amplicons. To compare the compositions of gut microbiota in children and adults of the same sex, a microbiota maturity index was created. In girls, similarity to adult-female
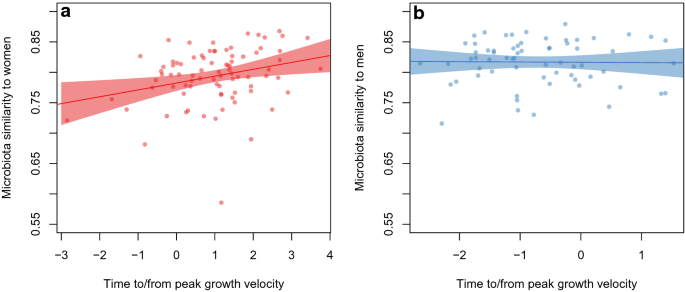
microbiota was positively correlated with pubertal progress: the more progressed the girl was in terms of puberty, the more similar were the microbiota (Fig. 1a, p = 0.009). The same was not
evident as regards boys versus adult males (Fig. 1b, p = 0.9). The association between the time from growth take-off and microbiota maturity was similar to that of the time from peak growth
velocity to maturity of the microbiota (girls, p = 0.01; boys, p = 0.85; Suppl. Figure 3). We tested whether antibiotic exposure, BMI, intake of probiotics, and intake of whole-grain foods
(based on dietary questionnaires) could explain the association between pubertal timing and the overall similarity to microbiota in adults. However, after adjustment for each of these
variables, the association remained significant in girls (p < 0.05). To assess the information on growth take-off and peak growth velocity, the children were divided into groups: prior to
growth take-off (N = 18/0 boys/girls), prior to peak growth (post take-off, up to three months before peak, N = 27/11 boys/girls), peak growth (three months before to six months after peak,
N = 13/17 boys/girls), immediately after peak growth (six months to two years after peak, N = 8/40 boys/girls) and more than two years after peak growth (N = 0/15 boys/girls). Principal
coordinates analysis revealed that these groups differed in microbiota composition in both sexes, with the composition becoming more adult-like as puberty progressed, especially in girls
(Fig. 2a). In girls, the summed score of the first two principal coordinates in the two post-peak groups was significantly higher than the score in the pre-peak group (< 2 years
post-peak, p = 0.02; > 2 years post-peak, p = 0.0004). In boys, the same trend was evident (Fig. 2b), although nonsignificant (compared with pre-take-off: pre-peak, p = 0.09; peak, p =
0.06; < 2 years post-peak, p = 0.07). The main driver behind maturation of the microbiota in girls was a change in the relative abundances of the two most abundant classes of bacteria,
Clostridiales and Bacteroidales (Fig. 3a–c). During puberty, the relative abundance of Clostridiales increased (p = 0.03, after adjusting for BMI, whole-grain intake, probiotic use and
antibiotic exposure) and the relative abundance of Bacteroidales decreased (p = 0.03) towards adult-like levels. The same trends, although nonsignificant (Clostridiales, p = 0.14;
Bacteroidales, p = 0.48), were evident in boys (Fig. 3d–f). At phylum level, the abundance of the Firmicutes increased with puberty progression in both sexes, significantly in girls (p =
0.01) and nonsignificantly in boys (p = 0.23), and the abundance of Bacteroidetes decreased (girls, p = 0.03; boys, p = 0.48). At family and genus levels, the puberty-associated changes in
the microbiota of girls were mainly attributable to an increase in members of Ruminococcaceae (_Anaerofilum_, p = 0.02; _Anaerotruncus_, p < 0.0001; _Subdoligranulum_, p = 0.02;
uncultured, p = 0.002) and _Lachnospiraceae_ (_Dorea_, p = 0.03; _Syntrophococcus_, p = 0.005) and a decrease of the order Bacteroidales (p = 0.03), especially _Paludibacter_ (p = 0.04),
_Macellibacteroides_ (p = 0.005) and _Barnesiella_ (p < 0.001). The same patterns were evident in boys, with only _Barnesiella_ (p < 0.001), _Macellibacteroides_ (p < 0.001) and
uncultured _Ruminococcaceae_ (p = 0.02) reaching statistical significance. In addition to the abundant taxa, there was a decline in low-abundance taxa _Streptococcus_ (girls, p = 0.02; boys,
p = 0.6), _Lactobacillu_s (girls, p = 0.5; boys, p = 0.009), _Coriobacteriaceae_ (girls, p = 0.1; boys, p = 0.04), and _Escherichia_ (girls, p = 0.3; boys, p < 0.001). We considered the
possibility that antibiotic exposure might influence the timing of puberty. For each sex, we identified the antibiotic type and timing of exposure that was most strongly associated with
pubertal timing, using AIC-based model selection. For girls, the most strongly associated antibiotic variable was (first- and second-generation) cephalosporin exposure (cumulative defined
daily doses, ddd) up to the age of 10 years, which was positively associated with pubertal progress (p = 0.004, Fig. 4a). In boys, cephalosporin exposure was not associated with the timing
of puberty (p = 0.97, Fig. 4b). For boys, the most strongly associated variable was macrolide exposure by the age of 10 years, which showed a non-significant negative trend (p = 0.12, Fig.
4d). In girls, macrolide use was not associated with puberty (p = 0.66, Fig. 4c). We also tested for an association between BMI and pubertal timing. In both sexes, there was a modest but
nonsignificant, positive trend (Suppl. Figure 4). Finally, we created multivariate models for the timing of puberty, including gut microbiota at family level, detailed information on
antibiotic exposure (time since last antibiotic course per antibiotic type, cumulative lifetime number of courses and ddd per antibiotic type), probiotic use, BMI, and gastrointestinal (GI)
symptoms as potential confounders and explanatory variables for both the timing of puberty and for the relative abundances of the bacterial taxa. We used AIC-based model selection to arrive
at the final model. Among girls, pubertal timing was associated positively with the abundance of Clostridiales family XIII bacteria and _Ruminococcace_ae, both members of the Clostridia
class (Fig. 5a). In addition, in a multivariate model, timing of puberty was associated positively with lifetime exposure to cephalosporins and negatively with self-reported flatulence. In
boys, a very different picture emerged. Timing of puberty was associated negatively with _Lactobacillaceae_ and _Pasteurellaceae_ and positively with _Neisseriaceae_ (Fig. 5b). The latter
taxa were associated with antibiotic exposure and with GI symptoms. In boys, the negative association between puberty and flatulence appeared to be mediated by _Neisseriaceae_. DISCUSSION In
this work we aimed to investigate the relationship between the timing of puberty and gut microbiota. Our results show, for the first time, that gut microbiota shifts towards adult-like
composition as the puberty progress. These findings suggest an association between sex hormones and gut microbiota development. Pubertal timing in girls was associated with antibiotic use,
especially cephalosporin. It is already known that antibiotic exposure, very likely via changes in the gut microbiota, is connected to higher BMI46,47, and higher BMI is connected to early
puberty48. However, in our dataset BMI was not significantly associated with pubertal timing and the association with cephalosporin use was also observable when adjusting for BMI as a
confounding factor. This suggests that antibiotic-driven changes in gut microbiota may be causally related to pubertal timing, independently of the metabolic effects. However, the connection
was not visible in boys in our cohort. Comparison of pubertal and adult microbiota revealed that during puberty, girls’ gut microbiota develops in the direction of adult women’s microbiota,
and the level of similarity is related to the degree of pubertal development. Importantly, the trend towards adult-like microbiota was visible in the first two principal coordinates,
indicating that the main dimensions of variation in the gut microbiota are affected by pubertal progression in this age group. Among boys a similar trend was observed with respect to the
first two principal coordinates (together explaining 32% of total microbiota variation), but not when considering the whole microbiota composition. It is possible that the route of
microbiota development is different in men compared with women, but the most probable explanation for the difference is the lack of data from boys past the period of peak growth velocity, as
a result of the generally later onset of puberty in boys, resulting in too few cases in this time window. For both sexes, the dominant taxa Clostridia, including the abundant families
_Ruminococcaceae_ and _Lachnospiraceae_, and Bacteroidia, including the abundant genus _Bacteroides_, and their ratio, approached adult levels in parallel with pubertal development. The
result shows that development of the gut microbiota that begins at the period of weaning49, with increasing abundance of Clostridia, still continues through puberty. Our data show that
children do not achieve an adult-like gut microbiota composition before the later stages of puberty. The increasing relative abundance of Clostridia with pubertal development is supported by
earlier findings. An Italian study revealed a lower abundance of Bacteroidetes and a higher abundance of Firmicutes in normal-weight adults compared with normal-weight adolescents, but the
pattern was opposite in obese individuals 29. In contrast, American study reported a higher abundance of Bacteroidetes in adults compared with 7- to 12-year-olds 28, corresponding to the
pattern seen in obese Italians. The weight status of the participants in the American study was not reported. A Dutch study in which 6- to 9-year-old children were compared with adults
showed a lower abundance of _Bacteroides_ and higher abundances of _Ruminococcus_, _Eubacterium_ and _Clostridium_ species in adults50. We had mostly normal-weight individuals, with only
four obese girls and five obese boys. Owing to small numbers it was not possible to reliably address whether or not gut microbiota development was different between obese and normal-weight
adolescents, but adjusting for BMI in the models did not alter the results. It is possible that obesity or, for example, a high-fat diet could alter microbiota progression during puberty. In
contrast to our findings, a recent Chinese study showed a higher abundance of Clostridia in 5-year-olds compared with 15-year-olds51. In the 15-year-olds, several genera were found to be
associated with the level of serum testosterone, but estradiol concentrations were not associated with microbiota composition51. Previous studies on the association between gut microbiota
and reproductive stage in humans have shown that at the end of female reproductive life, the pattern that we observed reverses again, with increasing abundance of Bacteroidetes and
decreasing abundances of Firmicutes during menopause52. Together, these results strongly indicate that sex hormones are a major driver of gut microbiota composition, particularly affecting
the balance between the two major phyla, Firmicutes and Bacteroidetes. Furthermore, _Bacteroides_ spp. were positively and _Ruminococcus_ spp. negatively correlated with testosterone levels
in a cohort of women with and without polycystic ovary syndrome53. Several _Ruminococcaceae_ OTUs (operational taxonomic units) were at a lower abundance in women with polycystic ovary
syndrome53, suggesting that their abundance is dependent on normal gonadal function of the host. Several types of Clostridia, especially species of the _Ruminococcaceae_ genera
_Faecalibacterium_ and _Ruminococcus_, are known to reactivate inactive conjugated forms of estrogen via their beta-glucuronidase activity13. The beta-glucuronidase enzymes of _Ruminococcus_
and _Faecalibacterium_ spp. have the ability to cleave both estrone and estradiol, while the enzyme of _Bacteroides fragilis_ shows catalytic activity only to estrone—orders of magnitude
weaker than that of the Clostridial enzymes13. The relative abundance levels of members of the _Ruminococcaceae_ family have been positively correlated with levels of circulating
estrogens14. In accordance with the higher estrogen-metabolizing activity of _Ruminococcus_ spp., the ratio of estrogen metabolites to estrogen parent compounds in urine has been shown to
correlate positively with the relative abundance of _Ruminococcus_ and negatively with that of _Bacteroides_ spp.15. Based on these data and our current results, it is plausible that gut
microbiota may partly regulate the onset of puberty and menopause via their estrogen metabolism. This hypothesis suggests that microbiota-targeting treatments may be effective in regulating
estrogen levels. Indeed, increased consumption of dietary fiber reduces the levels of circulating estrogens18. The effect has been shown to be caused specifically by wheat bran, but not by
oat or corn bran19. Notably, wheat-bran consumption decreases the relative abundance of _Ruminococcaceae_, which are specialized in resistant starch fermentation17. In addition to their
estrogen metabolism, Clostridia and Bacteroidia, the two dominant bacterial classes in the adult human gut, have widespread and varying effects on host physiology9. The families
_Lachnospiraceae_ and _Ruminococcaceae_ contain the most important butyrate producers, which are adapted to fermenting complex plant-based carbohydrates and are generally considered
important for a healthy gut9. Members of the _Ruminococcaceae_ family have been associated with healthy growth in a cohort of undernourished young children10. While the capacity of specific
gut microbes to metabolize estrogen suggests that gut microbiota may regulate puberty, the reverse is also possible. Sex hormones might affect maturation of the microbiota by directly
affecting the growth of specific taxa, by influencing immune reactions to gut microbes, by affecting bile-acid composition20, or in some as yet uncharacterized manner. More investigation is
warranted to better understand the underlining mechanisms. There are some limitations in our study. Our investigation did not include hormone level measurements, and so the pubertal timing
is based on the changes in growth velocity only. For boys 13 years-of-age was probably not the optimal time point to investigate puberty-related changes in gut microbiota, likely resulting
in weaker results than in the girls. The sample size in this study was substantially smaller than the whole 13-year follow-up sample. This was due to lack of growth and microbial data. The
main reason for the limited availability of the growth data is, that some of the participants have moved away from the capital-city region of Finland, and therefore we couldn’t attain the
growth information from the school health-service records of the local municipalities. In addition, many participants were unwilling to provide faecal samples, which contributed to the
reduced sample size. However, it is unlikely that willingness to provide a faecal sample or continued residence in the capital region would confound the association between puberty and gut
microbiota. Our study population was selected on the basis of its high risk of allergic disease—they have at least one parent with atopic disease. The population was collected from a
relatively limited geographic area in the capital-city region of Finland (population 1.5 million inhabitants and population density 800–3000 inhabitants/km2), 50% living in urban, 40%
suburban and 10% in rural areas. These factors may affect the generalization of our findings. CONCLUSION Previously very little was known about the development of human gut microbiota in
puberty. Our investigation opens interesting insights to the connection of the gut microbiota and pubertal development. Our data shows, that the development is sex-specific. Gut microbiota
were connected to pubertal timing on girls, but not on boys, possible due to later development pattern on boys. The timing of puberty may be affected by the gut microbiota, particularly
members of _Ruminococcaceae_, which may affect the timing via regulating host sex-hormone levels. Although we didn’t have hormonal data, growth curve analysis is a valid method to estimate
the timing of puberty, even in the absence of serial measurements of gonadotropin and sex steroid levels. Pubertal timing on girls correlates with prepubertal antibiotic usage. The effect of
lifestyle factors on pubertal timing may be mediated via changes in gut microbiota. More investigation is warranted to evaluate the generalizability of the findings and to clarify the
cause-and-effect relations. ETHICS APPROVAL AND CONSENT TO PARTICIPATE All experiments were performed in accordance with relevant guidelines and regulations. Written informed consent was
obtained from all subjects’ parents and/or legal guardians. The study was approved by the Helsinki University Hospital Ethics Committee. The ethical statement number is 78/13/03/03/2013.
DATA AVAILABILITY The raw dataset analysed during the current study is available in the European Nucleotide Archive (ENA) at EMBL-EBI under accession number PRJEB44673
(https://www.ebi.ac.uk/ena/browser/view/PRJEB44673). REFERENCES * Day, F. R. _et al._ Genomic analyses identify hundreds of variants associated with age at menarche and support a role for
puberty timing in cancer risk. _Nat. Genet._ 49(6), 6. https://doi.org/10.1038/ng.3841 (2017). Article CAS Google Scholar * Ullah, R. _et al._ Postnatal feeding with a fat rich diet
induces precocious puberty independent of body weight, body fat, and leptin levels in female mice. _Front. Endocrinol_ 10, 758. https://doi.org/10.3389/fendo.2019.00758 (2019). Article
Google Scholar * Kapczuk, K. Elite athletes and pubertal delay. _Minerva Pediatr._ 69(5), 415–426. https://doi.org/10.23736/S0026-4946.17.05044-7 (2017). Article PubMed Google Scholar *
Aksglaede, L., Juul, A., Olsen, L. W. & Sørensen, T. I. A. Age at puberty and the emerging obesity epidemic. _PLoS ONE_ 4(12), e8450. https://doi.org/10.1371/journal.pone.0008450 (2009).
Article ADS CAS PubMed PubMed Central Google Scholar * Deng, X. _et al._ Association between small fetuses and puberty timing: a systematic review and meta-analysis. _Int. J. Environ.
Res. Public Health_ 14(11), 1. https://doi.org/10.3390/ijerph14111377 (2017). Article Google Scholar * Li, X. F. _et al._ Role of the posterodorsal medial amygdala in predator odour
stress-induced puberty delay in female rats. _J. Neuroendocrinol._ 31(6), 1. https://doi.org/10.1111/jne.12719 (2019). Article CAS Google Scholar * Kao, K. T., Denker, M., Zacharin, M.
& Wong, S. C. Pubertal abnormalities in adolescents with chronic disease. _Best Pract. Res. Clin. Endocrinol. Metab._ 33(3), 101275. https://doi.org/10.1016/j.beem.2019.04.009 (2019).
Article CAS PubMed Google Scholar * Yan, J. _et al._ Gut microbiota induce IGF-1 and promote bone formation and growth. _PNAS_ 113(47), E7554–E7563.
https://doi.org/10.1073/pnas.1607235113 (2016). Article CAS PubMed PubMed Central Google Scholar * Korpela, K. Diet, microbiota, and metabolic health: trade-off between saccharolytic
and proteolytic fermentation. _Annu. Rev. Food Sci. Technol._ 9, 65–84. https://doi.org/10.1146/annurev-food-030117-012830 (2018). Article CAS PubMed Google Scholar * Blanton, L. V. _et
al._ Gut bacteria that prevent growth impairments transmitted by microbiota from malnourished children. _Science_ 351(6275), 1. https://doi.org/10.1126/science.aad3311 (2016). Article CAS
Google Scholar * Korpela, K. _et al._ Childhood BMI in relation to microbiota in infancy and lifetime antibiotic use. _Microbiome_ 5(1), 26. https://doi.org/10.1186/s40168-017-0245-y
(2017). Article CAS PubMed PubMed Central Google Scholar * Angelakis, E. Weight gain by gut microbiota manipulation in productive animals. _Microb. Pathog._ 106, 162–170.
https://doi.org/10.1016/j.micpath.2016.11.002 (2017). Article PubMed Google Scholar * Ervin, S. M. _et al._ Gut microbial β-glucuronidases reactivate estrogens as components of the
estrobolome that reactivate estrogens. _J. Biol. Chem._ 294(49), 18586–18599. https://doi.org/10.1074/jbc.RA119.010950 (2019). Article PubMed PubMed Central Google Scholar * Flores, R.
_et al._ Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: a cross-sectional study. _J. Transl. Med._ 10, 253. https://doi.org/10.1186/1479-5876-10-253
(2012). Article CAS PubMed PubMed Central Google Scholar * Fuhrman, B. J. _et al._ Associations of the fecal microbiome with urinary estrogens and estrogen metabolites in postmenopausal
women. _J. Clin. Endocrinol. Metab._ 99(12), 4632–4640. https://doi.org/10.1210/jc.2014-2222 (2014). Article CAS PubMed PubMed Central Google Scholar * Baker, J. M., Al-Nakkash, L.
& Herbst-Kralovetz, M. M. Estrogen–gut microbiome axis: Physiological and clinical implications. _Maturitas_ 103, 45–53. https://doi.org/10.1016/j.maturitas.2017.06.025 (2017). Article
CAS PubMed Google Scholar * Salonen, A. _et al._ Impact of diet and individual variation on intestinal microbiota composition and fermentation products in obese men. _ISME J._ 8(11),
2218–2230. https://doi.org/10.1038/ismej.2014.63 (2014). Article CAS PubMed PubMed Central Google Scholar * Goldin, B. R. _et al._ The effect of dietary fat and fiber on serum estrogen
concentrations in premenopausal women under controlled dietary conditions. _Cancer_ 74(3 Suppl), 1125–1131.
https://doi.org/10.1002/1097-0142(19940801)74:3+%3c1125::aid-cncr2820741521%3e3.0.co;2-5 (1994). Article CAS PubMed Google Scholar * Rose, D. P., Goldman, M., Connolly, J. M. &
Strong, L. E. High-fiber diet reduces serum estrogen concentrations in premenopausal women. _Am. J. Clin. Nutr._ 54(3), 520–525. https://doi.org/10.1093/ajcn/54.3.520 (1991). Article CAS
PubMed Google Scholar * Org, E. _et al._ Sex differences and hormonal effects on gut microbiota composition in mice. _Gut Microbes_ 7(4), 313–322.
https://doi.org/10.1080/19490976.2016.1203502 (2016). Article CAS PubMed PubMed Central Google Scholar * Yurkovetskiy, L. _et al._ Gender bias in autoimmunity is influenced by
microbiota. _Immunity_ 39(2), 400–412. https://doi.org/10.1016/j.immuni.2013.08.013 (2013). Article CAS PubMed Google Scholar * Markle, J. G. M. _et al._ Sex differences in the gut
microbiome drive hormone-dependent regulation of autoimmunity. _Science_ 339(6123), 1084–1088. https://doi.org/10.1126/science.1233521 (2013). Article ADS CAS PubMed Google Scholar *
Guo, Y. _et al._ Association between polycystic ovary syndrome and gut microbiota. _PLoS ONE_ 11(4), e0153196. https://doi.org/10.1371/journal.pone.0153196 (2016). Article CAS PubMed
PubMed Central Google Scholar * Dong, G. _et al._ The association of gut microbiota with idiopathic central precocious puberty in girls. _Front. Endocrinol._ 10, 1.
https://doi.org/10.3389/fendo.2019.00941 (2019). Article Google Scholar * Yuan, X., Chen, R., Zhang, Y., Lin, X. & Yang, X. Sexual dimorphism of gut microbiota at different pubertal
status. _Microb. Cell Fact_ 19(1), 152. https://doi.org/10.1186/s12934-020-01412-2 (2020). Article CAS PubMed PubMed Central Google Scholar * Agans, R. _et al._ Distal gut microbiota of
adolescent children is different from that of adults. _FEMS Microbiol. Ecol._ 77(2), 404–412. https://doi.org/10.1111/j.1574-6941.2011.01120.x (2011). Article CAS PubMed Google Scholar
* Ringel-Kulka, T. _et al._ Intestinal microbiota in healthy US young children and adults: a high throughput microarray analysis. _PLoS ONE_ 8(5), 64315.
https://doi.org/10.1371/journal.pone.0064315 (2013). Article ADS CAS Google Scholar * Hollister, E. B. _et al._ Structure and function of the healthy pre-adolescent pediatric gut
microbiome. _Microbiome_ 3(1), 36. https://doi.org/10.1186/s40168-015-0101-x (2015). Article PubMed PubMed Central Google Scholar * Del Chierico, F. _et al._ Gut microbiota markers in
obese adolescent and adult patients: age-dependent differential patterns. _Front. Microbiol._ 9, 1210. https://doi.org/10.3389/fmicb.2018.01210 (2018). Article PubMed PubMed Central
Google Scholar * Kukkonen, K. _et al._ Probiotics and prebiotic galacto-oligosaccharides in the prevention of allergic diseases: a randomized, double-blind, placebo-controlled trial. _J.
Allergy Clin. Immunol._ 119(1), 192–198 (2007). Article CAS PubMed Google Scholar * Kuitunen, M. _et al._ Probiotics prevent IgE-associated allergy until age 5 years in
cesarean-delivered children but not in the total cohort. _J. Allergy Clin. Immunol._ 123(2), 335–341. https://doi.org/10.1016/j.jaci.2008.11.019 (2009). Article PubMed Google Scholar *
Kallio, S., Kukkonen, A. K., Savilahti, E. & Kuitunen, M. Perinatal probiotic intervention prevented allergic disease in a Caesarean-delivered subgroup at 13-year follow-up. _Clin. Exp.
Allergy_ 49(4), 506–515. https://doi.org/10.1111/cea.13321 (2019). Article CAS PubMed Google Scholar * Korpela, K. _et al._ Cohort profile: Finnish Health and Early Life Microbiota
(HELMi) longitudinal birth cohort. _BMJ Open_ 9(6), e028500. https://doi.org/10.1136/bmjopen-2018-028500 (2019). Article PubMed PubMed Central Google Scholar * Rikken, B. & Wit, J.
M. Prepubertal height velocity references over a wide age range. _Arch. Dis. Child_ 67(10), 1277–1280 (1992). Article CAS PubMed PubMed Central Google Scholar * Salonen, A. _et al._
Comparative analysis of fecal DNA extraction methods with phylogenetic microarray: effective recovery of bacterial and archaeal DNA using mechanical cell lysis. _J. Microbiol. Methods_
81(2), 127–134. https://doi.org/10.1016/j.mimet.2010.02.007 (2010). Article CAS PubMed Google Scholar * Korpela, K. _et al._ Probiotic supplementation restores normal microbiota
composition and function in antibiotic-treated and in caesarean-born infants. _Microbiome_ 6, 1. https://doi.org/10.1186/s40168-018-0567-4 (2018). Article Google Scholar * Korpela, Katri,
_mare: Microbiota Analysis in R Easily. R package version 1.0._ 2016. [Online]. Available: https://github.com/katrikorpela/mare * Edgar, R. C. Search and clustering orders of magnitude
faster than BLAST. _Bioinformatics_ 26(19), 2460–2461. https://doi.org/10.1093/bioinformatics/btq461 (2010). Article CAS PubMed Google Scholar * Quast, C. _et al._ The SILVA ribosomal
RNA gene database project: improved data processing and web-based tools. _Nucleic Acids Res_ 41(D1), D590–D596. https://doi.org/10.1093/nar/gks1219 (2013). Article MathSciNet CAS PubMed
Google Scholar * R Core Team, _R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria._ 2019. [Online]. Available:
https://www.R-project.org/ * Jari Oksanen _et al._, _vegan: Community Ecology Package. R package version 2.5-4._ 2019. [Online]. Available: https://CRAN.R-project.org/package=vegan *
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D. & R Core Team, __nlme: Linear and Nonlinear Mixed Effects Models_. R package version 3.1-137_. 2018. [Online]. Available:
https://CRAN.R-project.org/package=nlme * Venables, W. N., & Ripley, B. D., _Modern Applied Statistics with S. Fourth Edition. Springer, New York_. ISBN 0-387-95457-0 (2002). * Wickham,
H., _ggplot2: Elegant Graphics for Data Analysis_. Springer-Verlag New York, 2016. [Online]. Available: https://ggplot2.tidyverse.org * _Microsoft Powerpoint for Mac_. 2020. [Online].
Available: https://www.microsoft.com * Stark, C. M., Susi, A., Emerick, J. & Nylund, C. M. Antibiotic and acid-suppression medications during early childhood are associated with obesity.
_Gut_ 68(1), 62–69. https://doi.org/10.1136/gutjnl-2017-314971 (2019). Article CAS PubMed Google Scholar * Block, J. P. _et al._ Early antibiotic exposure and weight outcomes in young
children. _Pediatrics_ 142(6), 1. https://doi.org/10.1542/peds.2018-0290 (2018). Article Google Scholar * Chen, L.-K. _et al._ Trajectory of body mass index from ages 2 to 7 years and age
at peak height velocity in boys and girls. _J. Pediatr._ https://doi.org/10.1016/j.jpeds.2020.11.047 (2020). Article PubMed PubMed Central Google Scholar * Korpela, K. & de Vos, W.
M. Early life colonization of the human gut: microbes matter everywhere. _Curr. Opin. Microbiol._ 44, 70–78. https://doi.org/10.1016/j.mib.2018.06.003 (2018). Article PubMed Google Scholar
* Zhong, H. _et al._ Impact of early events and lifestyle on the gut microbiota and metabolic phenotypes in young school-age children. _Microbiome_ 7(1), 2.
https://doi.org/10.1186/s40168-018-0608-z (2019). Article PubMed PubMed Central Google Scholar * Yuan, X., Chen, R., Zhang, Y., Lin, X. & Yang, X. Gut microbiota: effect of pubertal
status. _BMC Microbiol_ 20, 1. https://doi.org/10.1186/s12866-020-02021-0 (2020). Article CAS Google Scholar * Santos-Marcos, J. A. _et al._ Influence of gender and menopausal status on
gut microbiota. _Maturitas_ 116, 43–53. https://doi.org/10.1016/j.maturitas.2018.07.008 (2018). Article PubMed Google Scholar * Liu, R. _et al._ Dysbiosis of Gut microbiota associated
with clinical parameters in polycystic ovary syndrome. _Front. Microbiol._ 8, 1. https://doi.org/10.3389/fmicb.2017.00324 (2017). Article ADS Google Scholar Download references
ACKNOWLEDGEMENTS We thank the study participants and research nurse Rhea Paajanen, and Nick Bolton for language editing. FUNDING This study was partially supported by the Biocodex Microbiota
Foundation (AS), grant 1308255 (FINMIC) from the Academy of Finland (WMdV & MK), Foundation for Pediatric Research (SK, TR), Sigrid Juselius Foundation (TR) and the Spinoza Award 2008
of the Netherlands Organization for Scientific Research (WMdV). AUTHOR INFORMATION Author notes * These authors contributed equally: Katri Korpela and Sampo Kallio. * These authors jointly
supervised this work: Taneli Raivio and Mikael Kuitunen. AUTHORS AND AFFILIATIONS * Human Microbiome Research Program, Faculty of Medicine, University of Helsinki, Haartmaninkatu 3, P.O. Box
21, 00014, Helsinki, Finland Katri Korpela, Anne Salonen & Willem M. de Vos * New Childrenʼs Hospital, Pediatric Research Center, University of Helsinki and Helsinki University
Hospital, Stenbäckinkatu 9, P.O. Box 347, 00029, Helsinki, Finland Sampo Kallio, Matti Hero, Anna Kaarina Kukkonen, Päivi J. Miettinen, Erkki Savilahti, Ella Kohva, Laura Kariola, Maria
Suutela, Annika Tarkkanen, Taneli Raivio & Mikael Kuitunen * Laboratory of Microbiology, Wageningen University, Stippeneng 4, P.O. Box 8033, 6700 EH, Wageningen, The Netherlands Willem
M. de Vos * Translational Stem Cell Biology and Metabolism Research Program, Faculty of Medicine, University of Helsinki, Yliopistonkatu 3, P.O. Box 4, 00014, Helsinki, Finland Taneli Raivio
Authors * Katri Korpela View author publications You can also search for this author inPubMed Google Scholar * Sampo Kallio View author publications You can also search for this author
inPubMed Google Scholar * Anne Salonen View author publications You can also search for this author inPubMed Google Scholar * Matti Hero View author publications You can also search for this
author inPubMed Google Scholar * Anna Kaarina Kukkonen View author publications You can also search for this author inPubMed Google Scholar * Päivi J. Miettinen View author publications You
can also search for this author inPubMed Google Scholar * Erkki Savilahti View author publications You can also search for this author inPubMed Google Scholar * Ella Kohva View author
publications You can also search for this author inPubMed Google Scholar * Laura Kariola View author publications You can also search for this author inPubMed Google Scholar * Maria Suutela
View author publications You can also search for this author inPubMed Google Scholar * Annika Tarkkanen View author publications You can also search for this author inPubMed Google Scholar *
Willem M. de Vos View author publications You can also search for this author inPubMed Google Scholar * Taneli Raivio View author publications You can also search for this author inPubMed
Google Scholar * Mikael Kuitunen View author publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS K.K. and S.K. analyzed the data and were the major
contributors in writing of the manuscript. A.S. supervised and participated in the initial part of the data analysis. E.S., M.K., A.K. and S.K. collected and processed the Flora follow-up
data. T.R., M.H. and P.M. provided endocrinological expertize for the analysis and manuscript text. E.K., L.K., M.S. and A.T. analyzed the growth data. T.R. and M.K. did the initial
formulation of the investigation. W.M.dV., T.R. and M.K. had a major role in funding and supervision of the study. All authors contributed on the writing of the manuscript and read and
approved the final manuscript. CORRESPONDING AUTHOR Correspondence to Sampo Kallio. ETHICS DECLARATIONS COMPETING INTERESTS The authors declare no competing interests. ADDITIONAL INFORMATION
PUBLISHER'S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY FIGURE
LEGENDS. SUPPLEMENTARY FIGURE 1. SUPPLEMENTARY FIGURE 2. SUPPLEMENTARY FIGURE 3. SUPPLEMENTARY FIGURE 4. SUPPLEMENTARY TABLE 1. SUPPLEMENTARY TABLE 2. RIGHTS AND PERMISSIONS OPEN ACCESS This
article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as
you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party
material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the
article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the
copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Korpela, K., Kallio, S.,
Salonen, A. _et al._ Gut microbiota develop towards an adult profile in a sex-specific manner during puberty. _Sci Rep_ 11, 23297 (2021). https://doi.org/10.1038/s41598-021-02375-z Download
citation * Received: 02 June 2021 * Accepted: 12 November 2021 * Published: 02 December 2021 * DOI: https://doi.org/10.1038/s41598-021-02375-z SHARE THIS ARTICLE Anyone you share the
following link with will be able to read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer
Nature SharedIt content-sharing initiative