Response of soil bacteria on habitat-specialization and abundance gradient to different afforestation types
- Select a language for the TTS:
- UK English Female
- UK English Male
- US English Female
- US English Male
- Australian Female
- Australian Male
- Language selected: (auto detect) - EN
Play all audios:

ABSTRACT Studies involving response of subgroups of soil microorganisms to forest change, especially comparative studies on habitat-specialization and abundance gradient were still lack. In
this study, we analyzed the response of soil bacterial diversity and structure to afforestation types and its relationship to environment of Fanggan ecological restoration area under the
classification of subgroups by habitat-specialization and abundance gradient based on abundance ratio respectively. The results were: (1) On the habitat-specialization gradient, the
variation of OTUs species number and abundance was consistent and positively correlated with habitat-specialization; on the abundance gradient, the variation was opposite and OTUs species
number was negatively correlated with abundance gradient; (2) The distribution frequency of each subgroup on both gradients was the highest in broad-leaved forests, but the abundance was the
opposite. The distribution frequency of the same stand showed no difference among habitat-specialization subgroups, but the abundant subgroup in broad-leaved forests was the highest among
the abundance subgroups; (3) α-diversity was positively correlated with habitat-specialization but negatively with abundance, with the highest mostly in broad-leaved and mixed forests; (4)
Community structure among stands on habitat-specialization gradient showed no significant difference, but that of rare subgroup between broad-leaved forests and other stands significantly
differed. Plant diversity and vegetation composition correlated stronger with community structure than spatial distance and soil physicochemical properties on both gradients. Our results
provided a new perspective for revealing the effects of afforestation types on soil bacteria from the comparison of habitat specialization and abundance gradient. SIMILAR CONTENT BEING
VIEWED BY OTHERS SOIL HORIZONS REGULATE BACTERIAL COMMUNITY STRUCTURE AND FUNCTIONS IN DABIE MOUNTAIN OF THE EAST CHINA Article Open access 22 September 2023 SOIL MESO- AND MICRO-FAUNA
COMMUNITY IN RESPONSE TO BAMBOO-FUNGUS AGROFORESTRY MANAGEMENT Article Open access 30 September 2022 SOIL DEGRADATION INFLUENCES SOIL BACTERIAL AND FUNGAL COMMUNITY DIVERSITY IN OVERGRAZED
ALPINE MEADOWS OF THE QINGHAI-TIBET PLATEAU Article Open access 02 June 2021 INTRODUCTION Forest ecosystems played a crucial role in generating primary productivity, regulating nutrient
biogeochemical cycles and providing a variety of ecological services1. But with the rapid development of economy and society, a large number of natural forests have been deforested to meet
the demand for wood, which has been seriously damaged the forest ecosystems2,3. According to historical records, the primary forests in Shandong were almost cut down in the Qin and Han
dynasties, and the secondary forests were also destroyed repeatedly. Therefore, many areas of Shandong province had been bare rocks, and some had been artificial or secondary forests4.
Recent studies have shown that the barren hills in the northern Shandong central mountain area of began to suffering increasing forest cutting and land reclamation activities in the
fifteenth century5. During the sixteenth century to the eighteenth century, the local virgin forests were seriously destroyed or even cleared away, left barren mountains5. To alleviate the
crisis of the deforestation of natural forest, plantations have been rapidly rised and related researches have attracted the attention of researchers. However, plantations in Shandong
central mountain area began late, in the 1960s. Due to poor soil nutrients, conifers such as _Platycladus orientalis_ and _Pinus tabulaeformis_ were mainly planted6. Soil microorganisms are
key components that link the nutrient composition and cycle between above and below ground of forest ecosystem7,8. In recent years, interests in the factors that shape and control soil
microbial community composition and diversity have been spring up9, especially since the rise of high-throughput sequencing. Factors such as vegetation type, soil physicochemical properties,
plant diversity and spatial distance could affect the composition and structure of soil microbial community10,11,12. These factors interacted with each other, but their relative importance
to soil microbial diversity and community structure remains to be explored. Vegetation type was likely to be the main factor shaping soil microbial diversity and community structure13,14.
Different vegetation types affect soil microbial diversity and community structure through altering the chemical composition and content of litter and root exudates, making them strongly
related15,16. Vegetation types might also affect microbial community structure by regulating soil physicochemical properties17, in which soil pH, organic carbon could directly affect soil
microbial community composition18. Soil pH, carbon nitrogen ratio, etc., have also been shown vital for soil microbial community structures19,20. Studies have shown large differences in soil
microbial community structure between coniferous forests and broad-leaved forests21. In addition, high plant diversity contributed to the accumulation of soil organic matter and the
preservation of soil biodiversity22,23,24. Studies have also shown that diffusion being the main reason for the differences in soil microbial community structure25,26, this was often the
case in nutrient adequate ecosystems, because the impact of space distance on community structure was positively correlated with nutrient adequacy27. Relevant studies have mainly focused on
tropical forests22,28, subtropical forests23,29, temperate forests30and shrubs13, neglected the effect of afforestation types on soil microbial diversity and community structure of barren
hills in the north warm temperate zone. Although many factors have been proved to affect soil microbial diversity and community composition, different taxa responded different to
environmental factors. For example, fungal diversity was more sensitive to stand changes than bacteria31, while the latter was more sensitive to soil physicochemical properties32,33.
Different abundance and different habitat-specialization subgroups differed in their ability to adapt to and coexist with the environment, and their responses to soil environmental changes
were also different. Abundant subgroup tended to include fewer species and were widely distributed, and they tended to dominate community functions34. Rare subgroup tended to include more
species and more specialized distribution35,36. Previous studies have more focused to abundant subgroup which dominated community functions, because they often played a key role in organic
matter flow and biomass accumulation26, while rare subgroup has been ignored for a long time. However, the latter also played a key role in maintaining biodiversity and promoting nutrient
cycling and the function of diverse microbial communities37,38. On the abundance gradient, we defined the permanent abundant subgroup (relative abundance > 1% in all quadrats) and the
conditional abundant subgroup (relative abundance > 0.01% in all quadrats and > 1% in some quadrats) as abundant subgroup, the permanent rare subgroup (relative abundance < 1% in
all quadrats) and conditional rare subgroup (relative abundance < 0.01% in some quadrats and less than 1% in all quadrats) as rare subgroup and the rest were defined as medium abundance
subgroup39. On the gradient of habitat-specialization, the subgroup distributed evenly in each resource environment were classified into habitat-generalized subgroup. The poor niche
differentiation and severe uneven distribution caused by habitat changes were classified into habitat-specialized subgroup. The rest were habitat-neutral subgroup40. The classification of
habitat-specialization was based on the comparison between the actual niche width of OTUs and the confidence interval of zero niche width model. Those above the limit of 95% confidence
interval was classified into habitat-generalized subgroup, and those below the limit of 5% confidence interval was divided into habitat-specialized groups, and those between limit of 5 and
95% confidence interval was divided into habitat-neutral groups41. In this study, the responses of soil bacteria to different afforestation types were studied in a comparative way at both
habitat-specialization and abundance gradients, so as to reveal the differences and relationships of the above responses between abundance and niche differentiation. In the Fanggan
ecological restoration area located in the northern part of Shandong Central Mountain area, although there were plantations before the 1970s, the growth and forest formation effect was
little, and the restoration area was mostly Barren mountain and shrub (s) vegetation landform. Over the past 50 years, local villagers have moved 20,000,000 m3 of earth and planted more than
3,000,000 trees with different afforestation types, including coniferous forests (CF), mixed coniferous and broad-leaved forests (MF) and broad-leaved forests (BF). So far, the ecological
restoration area has formed the forest ecosystem with the coverage rate of more than 90% and a complete vertical community structure of tree, shrub and grass. At present, the restoration
area includes dense forest canopy zone, gaps with sparse forest canopy zone, dense shrub zone and bare rock lacking vegetation zone. The initial soil conditions of the restoration area were
consistent, so the current soil properties and microbial changes could reflect the effects of different afforestation types on soil characteristics. The results could not only reveal the
differences and mechanisms of response of soil bacterial to different afforestation types, but also provide valuable reference and guidance for Fanggan village to further strengthen and
adjust vegetation restoration strategies. MATERIALS AND METHODS STUDY SITE Fanggan ecological restoration area, locate in the northern part of Shandong Central Mountain area
(117°24′45″–117°28′5″E, 36°24′23″–36°26′44″N), was mountainous and hilly terrain with the highest altitude 860 m and more than 30 hilltops above 400 m. The climate type of the restoration
area is continental monsoon climate with distinct features of four seasons and the same period of rain and heat. The average annual temperature was 12.4 °C, the lowest extreme temperature
was − 22.5 °C and the average annual precipitation was more than 830 mm, concentrate in July to September. The soil type in the restoration area was mountain brown soil, and the average
thickness of litter was 4 cm. The vertical structure of vegetation community was obvious, but the species were relatively simple. The tree layer mainly included _Pinus densiflora_, _P.
tabulaeformis_, _Populus davidiana_, _Robinia pseudoacacia_ and _Diospyros lotus_, the shrub layer was mainly consisted of _Vitex negundo_ var. _heterophyll_, _Ziziphus jujuba_ var.
_spinosa_, _Grewia biloba_ and the regeneration seedlings of the tree layer, the herbaceous layer was consisted mainly of some species of Gramineae, Cyperaceae and Compositae. The
plantations in the restoration area were all planted between 1975 and 1985, with relatively consistent forest age. VEGETATION SURVEY AND SOIL SAMPLING Through literature review and field
investigation, forests of the restoration were mainly distributed in the altitude range of 300–750 m, so the vegetation community investigation was carried out in dense forest canopy region
of the altitude range. Because the original planting area of different forest types was quite different, this study allocated the quadrat quantity according to the actual area proportion of
each forest type. A total of 20 quadrats including 17 quadrats of 3 forest types of plantations, namely, 3 coniferous forests (CF), 3 coniferous broad-leaved mixed forests (MF) and 11
broadleaved forests (BF) as well as 3 shrubs quadrats as comparison were set up. In order to reduce the influence of topographic factors, each quadrat contained a consistent amplitude of
topographic variation as far as possible, and the geographical coordinates, elevation, slope direction and slope have been recorded (Table 1). The area of tree layer quadrat was 10 m × 10 m,
individuals with DBH ≥ 3 cm were measured per tree, and the species name, coverage, quantity, base diameter and height of shrubs as well as species name, height and coverage of herbs were
also recorded. The survey area of each shrub quadrats was 5 m × 5 m, and the statistics of herb layer were carried out. The α-diversity indexes calculated of vegetation community includes
Shannon-winner index (Eq. 1), Simpson diversity index (Eq. 2), Shannon evenness index (Eq. 3), Simpson evenness index (Eq. 4), Pielou evenness index (Eq. 5) and species richness. 0–10 cm
topsoil and 20–30 cm subsoil were collected in each quadrat according to the five-point sampling method with sampling points arranged at the center and four corners (3 m from the quadrat’s
vertex) of the quadrat35,42. The samples from same layer at 5 points in each quadrat were thoroughly mixed and divided into two portions. So, there were totally 80 soil sample, for one part
was stored in liquid nitrogen for molecular biology extraction and analysis, and the other was stored at room temperature and brought back to the laboratory for physicochemical analysis as
soon as possible. $${\text{Shannon-Winner}}\;{\text{diversity}}\;{\text{index}} = - \mathop \sum \limits_{i = 1}^{m} P_{i} \ln P_{i}$$ (1)
$${\text{Simpson}}\;{\text{diversity}}\;{\text{index}} = 1 - \mathop \sum \limits_{i = 1}^{m} \frac{{P_{i} \left( {P_{i} - 1} \right)}}{{N\left( {N - 1} \right)}}$$ (2)
$${\text{Shannon}}\;{\text{evenness}}\;{\text{index}} = \frac{Shannon - winner\;diversity\;index}{{{\text{log}}\left( m \right)}}$$ (3) $${\text{Simpson}}\;{\text{evenness}}\;{\text{index}}
= \frac{Simpson\;diversity\;index}{{{\text{log}}\left( m \right)}}$$ (4) $${\text{Pielou}}\;{\text{evenness}}\;{\text{index}} = \frac{{ - \mathop \sum \nolimits_{i = 1}^{m} P_{i} \log P_{i}
}}{{{\text{log}}\left( m \right)}}$$ (5) In the above equations, _m_ represents the number of species in the quadrat, _N_ represents the total number of individuals of all species, and _Pi_
represents the important value of species i in the quadrat. The diversity index of a stand is equal to the average value of the index for the various sides of the stand. DETERMINATION OF
SOIL PHYSICOCHEMICAL PROPERTIES Soil moisture and dry matter content were determined by gravimetric method43, pH was determined by potentiometric method using 2.5:1 water-soil ratio. Organic
carbon content was determined by potassium dichromate oxidation-spectrophotometry44, and available phosphorus was determined by molybdenum-antimony anti-color spectrophotometry45. We used
potassium chloride solution extraction-spectrophotometry and ultraviolet spectropotometry to determine ammonium nitrogen (NH4+-N) and nitrate nitrogen (NO3−-N) respectively. The content of
nitrite nitrogen (NO2−-N) was determined by naphthalene ethylenediamine hydrochloride color spectrophotometry46,47. The model of the spectrophotometer is T6 UV–visible spectrophotometer
produced by Beijing Puxi General Instrument Co., LTD. The pH meter model is SEvenExcellence S400-Basic made by Mettler Toledo Corporation. HIGH THROUGHPUT SEQUENCING OF SOIL MICROORGANISMS
Soil bacterial DNA was extracted by TGuide S96 magnetic bead method and 16S rDNA fragments of DNA were amplified by primers 515F (50-GTGYCAGCMGCCGCGGTAA-3) and 926R (50
-CCGYCAATTYMTTTRAGTTT-30). The PCR products were purified by the VAHTS-TM DNA Clean Beads method and quantified. Unique barcode was used to separate each sample to prevent
cross-contamination. Sequencing and data processing (1) Filter of raw data quality Trimmomatic48 was used to filter dual-end sequencing files. Parameter setting: Window size was set as 50
bp. The reads will be cut from the start of the window once average Q-score within the window is lower than 20. (2) Identification and removal of primer sequences Cutadapt (Version 1.9.1)
was used to identify primer sequences according to the parameters allowing the maximum error ratio of 20% and the minimum coverage of 80%49. (3) Double-ended reads splicing USEARCH (Version
10) was used for sample double-ends reads stitching according to the minimum overlap length of 10 bp, the minimum similarity of 90% allowed in overlap area and maximum error base number of 5
bp50. (4) Removal of chimera The criteria of chimera are as follows: divide query sequence into chunks without overlap and compare them with the database; The best match of each chunk in
the database is selected, and the two best parent sequences are finally selected. The sequence to be detected was compared with the two parents. If a sequence of the two parents has greater
than 80% similarity to the Query sequence, the query was judged to be a chimera. Use UCHIME (Version 8.1) to remove chimeras51. (5) OTUs clustering and species annotation Cluster analysis
was performed using UPARSE at the 97% similarity level, and the usearch command was used to remove affine sequences and singleton OTUs in the process. Each clustered OTUs was annotated into
seven subgroupomic classes of boundary, phylum, order, family, genus and species by the sine method through the 16 s in silva or the ITS database in Unite. The community abundance tables for
each rank were obtained. The present study was carried out mainly on the OTUs subgroupomic level and the following analysis were all based on the OTUs abundance table. DATA ANALYSIS
CLASSIFICATION ON ABUNDANCE GRADIENT In this study, to estimate the relative abundance of abundant and rare subgroups the thresholds were defined as 1 and 0.01% of the total sequence
reads52,53. Further to subdivide bacterial communities, the six categories were defined with reference to recent publications54,55: (1) permanent abundant subgroup (AAT): relative abundance
great than 1% in all samples, (2) permanent rare subgroup (ART): relative abundance less than 0.01% in all samples, (3) conditional medium abundance subgroup (MT): relative abundance between
0.01 and 1% in all samples, (4) conditionally abundant subgroup (CAT): relative abundance great than 0.01% in all samples and 1% in some samples, (5) conditionally rare subgroup (CRT):
relative abundance less than 0.01% in some samples but never great than 1% in any sample, (6) conditionally rare and abundant subgroup (CRAT): relative abundance ranging from 0.01% to 1%.
The above AAT and CAT were classified as abundant subgroup, ART and CRT as rare subgroup, and MT and CRAT as medium abundance subgroup. CLASSIFICATION ON HABITAT-SPECIALIZATION GRADIENTS
Levins niche width (Eq. 6) was calculated using the ‘spaa’ package of R v 4.0.056. The frequency of occurrence of each OTU was randomized and rearranged 1000 times using the replacement
method of the ‘EcolUtilis’ package, respectively. Then the zero distribution of the niche width index was calculated for each OTU. Based on the consensus that habitat-generalized subgroup
owe a wider niche width than the habitat-specialized subgroup40, OTUs were classified into a habitat-generalized or a specialized group based on whether the frequency of observed occurrence
exceeded the upper 95% confidence interval or fell below the lower 5% confidence interval, and the OTUs were distributed to the habitat-neutral group if the observed niche width was within
in the 5% to 95% confidence interval range18. $${\mathbf{B}}_{{\mathbf{i}}} = \frac{1}{{\mathop \sum \nolimits_{{{\varvec{j}} = 1}}^{{\varvec{r}}} {\varvec{P}}_{{{\varvec{ij}}}}^{2} }}$$ (6)
BACTERIAL COMMUNITY DIVERSITY CALCULATION Based on the OTUs abundance matrix, the Shannon-Winner diversity indexes and OTUs species richness of each habitat-specialization subgroups and
abundant subgroups were calculated. The non-metric multidimensional scaling (NMDS) function of ‘vegan’ package was used to analyze differences in bacterial subgroup community structure
between afforestation types, and the significance of structural differences was tested by Permanova test (by ANOSIM function in vegan package of R software, permutations = 999, bray). Mantel
test was carried out to analyze the relationship between community structure and environmental factors such as tree species composition, plant diversity, physicochemical properties and
spatial distance. The spatial distance was calculated by the three-dimensional relationship of latitude coordinates, longitude coordinates and altitude. RDA function was taken to analyze the
effects of tree composition, tree diversity and soil physicochemical properties on microbial subgroup community structure. The significance of RDA model was tested by ANOVA and the Monte
Carlo permutation test was taken to analyze the significance of the influencing factors of each explanatory variable in the RDA model (by anova.cca and envfit function in vegan package of R
software, permutations = 999). Plotting was done using the R 4.0.0 basic program package and ‘ggplot2’ package. The significance of soil physicochemical properties, niche width, abundance
and α-diversity indexes among forest types were analyzed by one-way ANOVA test (SPSS 19.0). RESULTS AND ANALYSIS A total of 2,506,761 sequences were tested from all soil samples, and were
divided into 1830 OTUs. The sequences number of 20 topsoil samples ranged from 46,595 to 73,284, and the OTUs ranged from 940 to 1579. Sequences number of 20 subsoil samples ranged from
47,705 to 70,743, and OTUs ranged from 1181 to 1568. On the habitat-specialization gradient, the OTUs numbers of habitat-generalized subgroup accounted for only 5.19%, and the abundance
ratio was only 1.70%. The habitat-specialized subgroup accounted for 65.41%, and the abundance ratio was also as high as 83.23% in topsoil. The OTUs number of the habitat-generalized
subgroup accounted for only 5.79% and the abundance ratio was only 1.83%, while the OTUs number of habitat-specialized subgroup was as high as 62.79% and the abundance ratio was as high as
81.88% in subsoil. On the abundance gradient, the OTUs numbers of abundant subgroup in the topsoil accounted for only 3.20%, but the abundance ratio was as high as 47.38%. OTUs numbers of
rare subgroup accounted for 93.82%, but the abundance ratio was only 41.21%. The OTUs numbers of abundant subgroup in subsoil only accounted for 1.87%, but the abundance ratio was as high as
36.55%. The OTUs numbers of rare subgroup accounted for as high as 94.84%, but the abundance ratio was only 47.22%. Soil physicochemical features were shown in Table 2. EFFECTS OF
AFFORESTATION TYPES ON THE DISTRIBUTION FREQUENCY AND ABUNDANCE OF SOIL BACTERIA ALONG HABITAT-SPECIALIZATION AND ABUNDANCE GRADIENTS Distribution frequency of soil bacteria was presented by
the niche width. Results showed the distribution frequency of each habitat-specialization subgroup in topsoil and subsoil was significantly the highest in broad-leaved forests (_P_ <
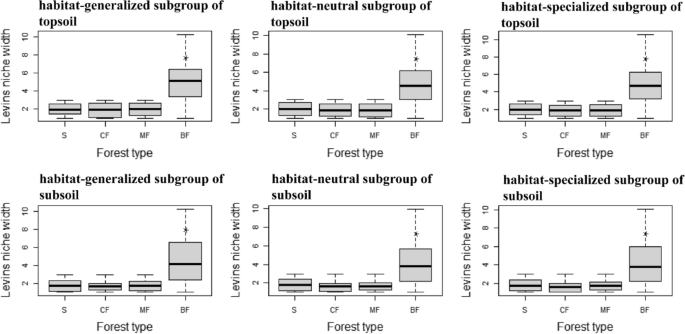
0.01), and even the frequency of habitat-neutral and specialized subgroups in coniferous forests and mixed forests was slightly lower than in the shrub (Fig. 1). The distribution frequency
of each abundance subgroup in topsoil and subsoil was also increased obviously in broad-leaved forests, especially for abundant subgroups (_P_ < 0.01). The niche width of the abundant
subgroup in broad-leaved forests also showed higher than other subgroups (Fig. 2). On the habitat-specialization gradient, the abundance of each subgroup in topsoil has increased with
habitat-specialization. The abundance of the same habitat-specialization subgroup in the topsoil and subsoil showed consistent with the stand changes. The abundance of habitat-generalized
and neutral subgroups was higher in shrub and mixed forests than in the coniferous forests and broad-leaved forests and that of habitat-specialized subgroup showed the highest in the shrub
and the lowest in the broad-leaved forests, and significantly differ (_P_ < 0.05) (Fig. 3). On the abundance gradient, the abundance of abundant and medium abundance subgroups in the
topsoil and subsoil both significantly decreased, whereas that of rare subgroup increased in broad-leaved forests. This indicated the soil conditions of broad-leaved forests were more
comfortable for most species of the rare subgroup. The abundance of abundant subgroup of both soil layer in coniferous forests and mixed forests were significantly higher than in shrub and
broad-leaved forests. The abundance of medium abundance subgroup was lower in the topsoil of broad-leaved forests than in other stands, but obviously the trend of shrub > coniferous
forests > mixed forests > broad-leaved forests in the subsoil. The abundance features of rare subgroup in topsoil were opposite to those of abundant subgroup (Fig. 4). EFFECTS OF
AFFORESTATION TYPES ON SOIL BACTERIAL Α-DIVERSITY CHARACTERISTICS ALONG HABITAT-SPECIALIZATION AND ABUNDANCE GRADIENTS Shannon-Winner index of habitat-generalized subgroup of each
afforestation stands in both topsoil and subsoil was significantly higher than that of shrub in all tree forests (_P_ < 0.05) (Fig. 5(TG)(SG)), but only the index of habitat-specialized
and neutral subgroups in subsoil of broad-leaved forests significantly higher than other stands (_P_ < 0.05) (Fig. 5(TS)(SS)), and the habitat-neutral subgroup of topsoil were almost
unaffected by afforestation (Fig. 5(TN)). The species richness of habitat-specialized subgroup in topsoil was significantly higher in broad-leaved forests than in other stands (_P_ <
0.01) (Fig. 5(TN)), while that of habitat-generalized and neutral subgroups was highest in mixed forests (Fig. 5(TG)(TN)). The species richness of each habitat-specialization subgroups in
subsoil was higher in broad-leaved forests and mixed forests than in coniferous forests and shrub (Fig. 5(SG)(SN)(SS)). The diversity of rare subgroup in both soil layer was higher than that
of abundant and medium abundance subgroups. Shannon-Winner index of abundant and rare subgroups in topsoil and subsoil was higher in broad-leaved forests and mixed forests than in
coniferous forests and shrub (Fig. 6(TA)(SA)(TR)(SR)). Species richness of topsoil rare subgroup was significantly higher in broad-leaved forests than in other stands, and that of rare
subgroup in subsoil was significantly higher in broad-leaved forests and mixed forests than in coniferous forests and shrub (_P_ < 0.05). Shannon-diversity index and species richness were
consistent among forest types in both topsoil and subsoil (Fig. 6(TM)(SM)). RESPONSES AND DRIVING FACTORS OF SOIL BACTERIAL COMPOSITION AND STRUCTURE TO AFFORESTATION TYPES ALONG
HABITAT-SPECIALIZATION AND ABUNDANCE GRADIENTS For OTUs with abundance greater than 1%, species number of habitat-generalized subgroup was significantly higher than that of habitat-neutral
and specialized subgroups, and the abundance distribution was more uniform in the topsoil (Fig. 7a, b, c). Among all forest types, the distribution uniformity of abundance ratio was the best
in broad-leaved forests. The abundance ratio of top 3 OTUs in topsoil habitat-generalized and neutral subgroups were the highest in the coniferous forests (Fig. 7a, b), while the highest of
specialized subgroup was in the shrub (Fig. 7a). The abundance ratio of each OTUs of all habitat-specialization subgroups in broad-leaved forests was the lowest, which was consistent with
its higher distribution uniformity. The abundance ratio of top 3 OTUs of habitat-generalized subgroup was the highest in shrub, and the highest of the habitat-neutral and specialized
subgroups was in the coniferous forests (Fig. 8a, b, c). Among the top three abundant subgroups of topsoil, the highest abundance ratio was found in the coniferous forests (Fig. 9a), the
highest abundance of medium abundance subgroup was found in the mixed forests and the highest in shrub of rare subgroup (Fig. 9b), and the highest abundance was found in the shrub layer in
the rare groups (Fig. 9c). In subsoil, the highest proportion of OTUs of abundant subgroup was found in mixed forests (Fig. 10a), and that of medium abundance and rare subgroups was found in
shrub (Fig. 10b, c). NMDS analysis was carried out on the community structural difference among afforestation types along habitat-specialization and abundance gradient. The 2D figures
showed that the coordinate points of the rare subgroup in both layers of the broad-leaved forests were significantly separated from other stands, that is, the coordinate points of the
broad-leaved forests were distributed at the upper right position of the figure, while that of other stands distributed at the lower left. The coordinate points of community structure of
each habitat-specialization subgroup could not be distinguished among each stand, and the same for abundant and medium abundance subgroups (Figs. 11, 12). However, in the right half of all
NMDS maps, only coordinate points of broad-leaved forests were distributed, indicating that in some broad-leaved forests, bacterial community structure had been significantly different from
that of other stands (Fig. 11). Further ANOSIM analysis showed that afforestation affected non-significantly on the community structure of each habitat-specialization subgroup in both
topsoil and subsoil, but significantly affected the community structure of the rare subgroup in topsoil. The environmental affection on the community structure among afforestation types of
each subgroup have also been analyzed. Mantel analysis showed that plant diversity and vegetation species composition rather than soil chemical properties and spatial distance significantly
affected bacterial community structure among habitat-specialization and abundant subgroups. Plant diversity significantly affected the structure of each habitat-specialization subgroup among
afforestation types in both layers, and the species composition of tree layer significantly affected the structure of each habitat-specialization subgroup in topsoil and habitat-specialized
subgroup in subsoil. The species composition of shrub layer significantly affected the structure of habitat-specialized subgroup in topsoil (Table 3). The species composition of tree layer
significantly affected the structure of all abundance subgroups except abundant subgroup of subsoil. Plant diversity and species composition of shrub layer significantly affected the
structure of medium abundance and rare subgroups in topsoil (Table 4). RDA ANALYSIS OF EFFECTS OF VEGETATION AND ENVIRONMENTAL FACTORS ON SOIL BACTERIAL STRUCTURE ALONG
HABITAT-SPECIALIZATION AND RICHNESS GRADIENTS EFFECT OF SPECIES COMPOSITION On the habitat-specialization gradient, the RDA model of whole species composition of tree layer significantly
explained the community structure of each habitat-specialization subgroup in both topsoil and subsoil. But the change of important value of any individual tree species did not show
significant effect in the topsoil. The importance values of _Robinia pseudoacacia_ and _Morus alba_ significantly affected the community structure of the habitat-specialized subgroup in
subsoil (Table S1, Fig. S1). The RDA model of species composition of shrub layer significantly explained the community structure of habitat-generalized and neutral subgroups in the subsoil.
Individual shrub species composition did not significantly affect the community structure of habitat-generalized subgroup in the topsoil. The significant impact factors for habitat-neutral
subgroup in topsoil was _Platycladus orientalis_, _Robinia pseudoacacia_ and _Morus mongolica_, for habitat-specialized subgroup in topsoil was _Vitex negundo_ var. _heterophylla_, for
habitat-generalized subgroup in subsoil was _Vitex negundo_ var. _heterophylla_ and _Vitex negundo_ var. _heterophylla_ and for habitat-neutral and specialized subgroups in subsoil was
_Robinia pseudoacacia_ and _Morus mongolica_ (Table S2, Fig. S2). On the abundance gradient, the RDA models of overall species composition of tree layer for explaining the community
structure of medium abundance and rare subgroups of topsoil as well as abundant and rare subgroups of subsoil were significant. In the topsoil, the significant impact species for abundant
subgroup were of _Quercus acutissima_ and _Pinus densiflora_, for medium abundance subgroup was _Populus davidiana_, _Diospyros lotus_ and for rare subgroup was _Robinia pseudoacacia_ and
_Populus davidiana_. In the subsoil, significant impact factors of abundant subgroup were _Platycladus orientalis_, _Robinia pseudoacacia_, _Morus alba_ and _Ziziphus mauritiana_, and for
medium abundance subgroup were _Robinia pseudoacacia_ and _Morus alba_ (Table S3, Fig. S3). The overall species composition of shrub layer could not significantly explain the changes of
community structure of each abundance subgroup. Monte Carlo analysis also showed that only _Vitex negundo_ var. _heterophylla_, _Vitex negundo_ var. _heterophylla_ and _Morus mongolica_
significantly affected the community structure of a few subgroups (Table S4, Fig. S4). EFFECT OF PLANT DIVERSITY The RDA model of plant diversity feature to explain the community structure
of each habitat-specialization subgroup in the topsoil and subsoil was significant. Monte Carlo test results has also showed that all the diversity indices significantly affected the
structure of each subgroup in the topsoil, and the evenness indices significantly affected the structure of each subgroup in the subsoil (Table S5, Fig. S5). For each abundance subgroup, all
diversity indexes have significantly affected the structure changes of rare subgroup in topsoil and medium abundance subgroups in subsoil. Shannon-Winner diversity index, Simpson diversity
index and Pielou evenness index had significant effects on the structure of other subgroups in topsoil and subsoil (Table S6, Fig. S6). EFFECT OF SOIL CHEMICAL PROPERTIES The RDA model of
soil physicochemical properties to explain the community structure of habitat-neutral and specialized subgroups in topsoil and habitat-generalized subgroup in subsoil was significant.
However, Monte Carlo test showed that the significant factors affecting the structure of each subgroup were different (Table S7). According to the RDA 2D figure, the differences of each
habitat-specialization subgroup in the topsoil has been more affected by soil physicochemical properties than that in the subsoil. The coordinates community structure of the
habitat-generalized and neutral subgroups of broad-leaved forests in topsoil directed the same as the increase of pH, available P and ammonia nitrogen, indicating that these physicochemical
properties have significantly changed the structure of the two subgroups of in the topsoil of broad-leaved forests, while the coordinate points of the habitat-specialized subgroup in subsoil
of broad-leaved forests were contrary to the changes of ammonia nitrogen and pH, indicating that ammonia nitrogen and pH also significantly affected the structure habitat-specialized
subgroup in subsoil. No significant difference has been proved in the structure of other subgroups among stands (Fig. S7). For each abundance subgroup, soil physicochemical properties
significantly explained the changes of community structure of rare subgroup in topsoil. Monte Carlo test showed the structure of abundant subgroup significantly correlated with the content
of available P and nitrate nitrogen, and that of medium abundance and rare subgroup significantly correlated with the content of ammonia nitrogen and pH. In the subsoil, the structure of
abundant subgroup was significantly correlated with ammonia nitrogen content, and that of medium abundance subgroup was significantly correlated with dry matter content and ammonia nitrogen
content, as well as the structure of rare subgroup was significantly correlated with dry matter, organic carbon, nitrate nitrogen content and pH (Table S8, Fig. S8). DISCUSSION RESPONSE OF
ABUNDANCE AND DISTRIBUTION FREQUENCY OF SOIL BACTERIA TO AFFORESTATION ALONG HABITAT-SPECIALIZATION AND ABUNDANCE GRADIENTS Research results have shown that the consistency of variation of
species number and abundance of soil bacterial OTUs were significantly different on the two gradients. At the habitat-specialization gradient, the species number and abundance ratio changed
consistently and showed the highest of habitat-specialized subgroup while the lowest of habitat-generalized subgroup. This was consistent with the research of Estavillo et al. about forest
loss and biodiversity threshold57. However, at the abundance gradient, the abundant subgroup owns a small number of OTUs species and a high abundance ratio, while the rare subgroup showed
the opposite. Some studies also showed that the total number of sequences of abundant subgroup was dominant while the species number of rare subgroup was absolutely dominant18. The
distribution frequency of soil bacterial OTUs was characterized by Levins niche width. The higher the value is, the more evenly distributed the individuals are in each resource state, that
is, the higher the degree of niche differentiation and the better the interspecies co-existence. However, a lower value means that the distribution of individuals in each resource state is
different and the co-existence between species is bad. Our results showed that the response of distribution frequency to stand change was highly consistent between the two subgroups
classification gradients. The broad-leaved forests significantly promoted the coexistence and niche differentiation of each habitat-specialization subgroup, and most positively promoted the
distribution frequency of abundant subgroup on the abundance gradient. The niche width of each habitat-specialization subgroup in the same forest type were similar, but that of the abundant
subgroup were higher than the medium abundance and rare subgroups, especially in the broad-leaved forests. This was because abundant subgroup existed more abundant-dominant OTUs than rare
subgroup, allowing them more competitive and thus adapt quickly to changing environments54. The response of bacterial OTUs abundance to stand changes also differ between the two gradients.
The abundance of each habitat-specialization subgroup was the lowest in the broad-leaved forests, which could be interpreted as that the soil environment of broad-leaved forests was
conducive to niche differentiation among species and reduced the occurrence probability of a few high-abundance species58. The abundance of abundant and medium abundance subgroups in
broad-leaved forests also decreased, but the abundance of rare subgroup increased, which could be interpreted as the decline in abundance of abundant subgroup enhanced the abundance of rare
subgroup and occupied new niches59,60. RESPONSE OF Α DIVERSITY CHARACTERISTICS TO AFFORESTATION ALONG HABITAT-SPECIALIZATION AND ABUNDANCE GRADIENTS The α diversity of habitat-specialized
subgroup and rare subgroup was the highest on their respective gradients. This was consistent with the conclusion reported by Wu et al. that the diversity and species richness of rare
subgroup in topsoil and subsoil were significantly higher than those of abundant and medium abundance subgroups58. In general, the diversity index of each subgroup in broadleaved forests was
higher than that of other stands61, and the response of diversity to stand change on habitat-specialization gradient was stronger than that on abundance gradient, of which the
habitat-specialized subgroup responded the strongest. A number of studies have shown that bacterial habitat specialists were more susceptible to extinction than bacterial habitat generalists
when habitat conditions changed62,63, and this could legitimately interpret the fact that bacterial α-diversity of habitat-specialization gradient more strongly responded to stand changes
than the abundance gradient. The abundance gradient only represents the proportion of individual number of species, and the response of species and individual numbers of each subgroup to
stand change were similar and intersecting, so the response of α diversity to stand change on the abundance gradient was weak. RESPONSES OF SOIL BACTERIAL COMPOSITION AND COMMUNITY STRUCTURE
TO AFFORESTATION ALONG HABITAT-SPECIALIZATION AND ABUNDANCE GRADIENTS The results showed that the response of the composition and distribution features of bacteria to stand change was
different between two classification gradients. In the broadleaved forests, the distribution uniformity of each habitat-specialization subgroup was the highest. However, on the abundance
gradient, the most uniform distribution of OTUs in the broad-leaved forests was only in the abundant and rare subgroup of topsoil and rare subgroup of subsoil. This was consistent with the
fact that rare subgroup owns a large number of species but a low abundance, which resulted in a more even distribution of abundance than the abundant and medium abundance subgroups64,65. In
addition, the most uniform distribution in broad-leaved forests and the worst distribution uniformity in shrub and coniferous forests, was consistent with the conclusion on the increasing
complexity of soil bacterial network from temperate coniferous forests succession to broad-leaved forests in Liupanshan Mountain66, which promoted co-existence and niche differentiation by
reducing intraspecific and interspecific competition of bacteria67. Coordinate points of community structure of each habitat-specialization subgroup of some broad-leaved forests were
obviously seperated from that of other stands. These indicated the soil environmental changes in broad-leaved forests lead to significant changes in soil bacterial community structure on the
gradient of habitat-specialization. This conclusion was consistent with that the distribution frequency of each habitat-specialization subgroup in broad-leaved forests significantly higher
than other stands thus the community structure must be obviously different from that of other stands in which a few OTUs dominated. In terms of abundance gradient, ANOSIM analysis showed
that the differences among each forest type only significantly affected the community structure of rare subgroup in the topsoil, because their niche width was narrow and more sensitive to
environmental changes68, while the abundant subgroup tended to be more resilient and robust to environmental changes. The conclusion of this study provides a scientific reference and a new
perspective for the comparative study of soil microbial response to stand change from the two gradients of soil microbial habitat specialization and abundance. DATA AVAILABILITY SRA:
https://www.ncbi.nlm.nih.gov/bioproject/PRJNA852687. REFERENCES * Sasmito, S. D. _et al._ Organic carbon burial and sources in soils of coastal mudflat and mangrove ecosystems. _CATENA_ 187,
104414. https://doi.org/10.1016/j.catena.2019.104414 (2020). Article CAS Google Scholar * Tian, D. _et al._ A long-term evaluation of biomass production in first and second rotations of
Chinese fir plantations at the same site. _Forestry_ 84, 411–418 (2011). Article Google Scholar * Jacobson, S. Addition of stabilized wood ashes to Swedish coniferous stands on mineral
soils-Effects on stem growth and needle nutrient concentrations. _Silva Fenn._ 37(4), 437–450 (2003). Article Google Scholar * Li, J. H. Historical evolution of forests in Shandong
Province Agricultural. _Archaeology_ 01, 219–225 (1987) (IN CHINESE). Google Scholar * Tan, L. C. _et al._ A multiple-proxy stalagmite record reveals historical deforestation in central
Shandong, northern China. _Sci. Sin._ 50(11), 1643–1654 (2020) (IN CHINESE). Google Scholar * Du, Z. Y. _et al._ Soil quality characteristics of _Platycladus orientalis_ plantations with
different densities in central mountainous area of Shandong Province. _J. Cent. South Univ. For. Technol._ 40(09), 104–112. https://doi.org/10.14067/j.cnki.1673-923x.2020.09.012 (2020) (IN
CHINESE). Article CAS Google Scholar * Jiao, S. _et al._ Soil microbiomes with distinct assemblies through vertical soil profiles drive the cycling of multiple nutrients in reforested
ecosystems. _Microbiome_ 6, 146. https://doi.org/10.1186/s40168-018-0526-0 (2018). Article PubMed PubMed Central Google Scholar * Wagg, C., Schlaeppi, K., Banerjee, S., Kuramae, E. E.
& van der Heijden, M. G. A. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. _Nat. Commun._ 10, 4841. https://doi.org/10.1038/s41467-019-12798-y
(2019). Article CAS ADS PubMed PubMed Central Google Scholar * Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. _FEMS Microbiol. Rev_ 41, 109–130.
https://doi.org/10.1093/femsre/fuw040 (2017). Article CAS PubMed Google Scholar * Yao, M. J. _et al._ The differentiation of soil prokaryote communities along a precipitation and
temperature gradient in the eastern Inner Mongolia steppe. _Catena_ 152, 47–56. https://doi.org/10.1016/j.catena.2017.01.00 (2017). Article CAS Google Scholar * Geml, J. _et al._ Changes
in richness and community composition of ectomycorrhizal fungi among altitudinal vegetation types on Mount Kinabalu in Borneo. _New Phytol._ 215, 454–468. https://doi.org/10.1111/nph.14566
(2017). Article CAS PubMed Google Scholar * Wu, B. _et al._ The biogeography of fungal communities in wetland sediments along the Changjiang River and other sites in China. _ISME J._ 7,
1299–1309. https://doi.org/10.1038/ismej201329 (2013). Article CAS PubMed PubMed Central Google Scholar * Canini, F. _et al._ Vegetation, pH and water content as main factors for
shaping fungal richness, community composition and functional guilds distribution in soils of western Greenland. _Front. Microbiol._ 110, e2348. https://doi.org/10.3389/fmicb.2019.02348
(2019). Article Google Scholar * Porazinska, D. L. _et al._ Plant diversity and density predict belowground diversity and function in an early successional alpine ecosystem. _Ecology_ 99,
1942–1952. https://doi.org/10.1002/ecy.2420 (2018). Article PubMed Google Scholar * Horrocks, C. A. _et al._ Smart forage selection could significantly improve soil health in the tropics.
_Sci. Total Environ._ 688, 609–621. https://doi.org/10.1016/j.scitotenv.2019.06.152 (2019). Article CAS ADS PubMed PubMed Central Google Scholar * Qu, Z. _et al._ The response of the
soil bacterial community and function to forest succession caused by forest disease. _Funct. Ecol._ 34, 2548–2559. https://doi.org/10.1111/1365-2435.13665 (2020). Article Google Scholar *
Yang, K. & Zhu, J. The effects of N and P additions on soil microbial properties in paired stands of temperate secondary forests and adjacent larch plantations in Northeast China. _Soil
Biol. Biochem._ 90, 80–86. https://doi.org/10.1016/j.soilbio.2015.08.002 (2015). Article CAS Google Scholar * Wu, W., Wang, X., Ren, Z., Zhou, X. & Du, G. N-induced species loss
dampened by clipping mainly through suppressing dominant species in an alpine meadow. _Front. Plant Sci._ 13, 815011. https://doi.org/10.3389/fpls.2022.815011 (2022). Article PubMed PubMed
Central Google Scholar * Zhalnina, K. _et al._ Soil pH determines microbial diversity and composition in the park grass experiment. _Microb. Ecol._ 69(2), 395–406 (2015). Article CAS
PubMed Google Scholar * Wan, X. H. _et al._ Soil C: N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations.
_Plant Soil_ 387(1–2), 103–116 (2015). Article CAS Google Scholar * Vuong, T. M. D., Zeng, J. Y. & Man, X. L. Soil fungal and bacterial communities in southern boreal forests of the
Greater Khingan Mountains and their relationship with soil properties. _Sci. Rep._ 10(1), 22025. https://doi.org/10.1038/s41598-020-79206-0 (2020). Article CAS ADS PubMed PubMed Central
Google Scholar * Lange, M. _et al._ Biotic and abiotic properties mediating plant diversity effects on soil microbial communities in an experimental grassland. _PLoS One_ 95, e96182.
https://doi.org/10.1371/journal.pone.0096182 (2014). Article CAS ADS Google Scholar * Lange, M. _et al._ Plant diversity increases soil microbial activity and soil carbon storage. _Nat.
Commun._ 6, 6707 (2015). Article CAS ADS PubMed Google Scholar * Wang, H. _et al._ Mixed-species plantation with _Pinus massoniana_ and _Castanopsis hystrix_ accelerates C loss in
recalcitrant coniferous litter but slows C loss in labile broadleaf litter in southern China. _For. Ecol. Manag._ 422, 207–213. https://doi.org/10.1016/j.foreco.2018.04.024 (2018). Article
Google Scholar * Wu, Y. C. _et al._ pH is the primary determinant of the bacterial community structure in agricultural soils impacted by polycyclic aromatic hydrocarbon pollution. _Sci.
Rep._ 7, 40093 (2017). Article CAS ADS PubMed PubMed Central Google Scholar * Liu, L. M., Yang, J., Yu, Z. & Wilkinson, D. M. The biogeography of abundant and rare bacterioplankton
in the lakes and reservoirs of China. _ISME J._ 9, 2068–2077. https://doi.org/10.1038/ismej.2015.29 (2015). Article PubMed PubMed Central Google Scholar * Gravel, D. _et al._
Reconciling niche and neutrality: The continuum hypothesis. _Ecol. Lett._ 9(4), 399–409. https://doi.org/10.1111/j.1461-0248.2006.00884.x (2006). Article PubMed Google Scholar * Mueller,
R. C. _et al._ Links between plant and fungal communities across a deforestation chronosequence in the Amazon rainforest. _ISME J._ 8, 1548–1550. https://doi.org/10.1038/ismej.2013.253
(2014). Article CAS PubMed PubMed Central Google Scholar * Han, W. _et al._ Effects of vegetation type, season, and soil properties on soil microbial community in subtropical forests.
_Appl. Soil Ecol._ 158, 103813. https://doi.org/10.1016/j.apsoil.2020.103813 (2021). Article Google Scholar * Nielsen, U. N., Osler, G. H. R., Campbell, C. D., Burslem, D. F. R. P. &
Wal, R. V. D. The influence of vegetation type, soil properties and precipitation on the composition of soil mite and microbial communities at the landscape scale. _J. Biogeogr._ 37,
1317–1328. https://doi.org/10.1111/j.1365-2699.2010.02281.x (2010). Article Google Scholar * Xu, H. D., Yu, M. K. & Cheng, X. R. Abundant fungal and rare bacterial taxa jointly reveal
soil nutrient cycling and multifunctionality in uneven-aged mixed plantations. _Ecol. Indic._ 129, 107932. https://doi.org/10.1016/j.ecolind.2021.107932 (2021). Article CAS Google Scholar
* Yin, Y., Li, Q. & Du, H. Near-natural transformation of _Pinus tabuliformis_ better improve soil nutrients and soil microbial community. _Peer J._ 9, e12098.
https://doi.org/10.7717/peerj.12098 (2021). Article PubMed PubMed Central Google Scholar * Han, S. _et al._ Bacterial rather than fungal diversity and community assembly drive soil
multifunctionality in a subtropical forest ecosystem. _Env. Microbiol. Rep._ 14(1), 85–95. https://doi.org/10.1111/1758-2229.13033 (2022). Article CAS ADS Google Scholar * Lynch, M. D.
J. & Neufeld, J. D. Ecology and exploration of the rare biosphere. _Nat. Rev. Microbiol._ 13, 217–229. https://doi.org/10.1038/nrmicro3400 (2015). Article CAS PubMed Google Scholar *
Pedrós-Alió, C. Marine microbial diversity: Can it be determined?. _Trends Microbiol._ 14(6), 257–263. https://doi.org/10.1016/j.tim.2006.04.007 (2006). Article CAS PubMed Google Scholar
* Logares, R., Mangot, J. F. & Massana, R. Rarity in aquatic microbes: placing protists on the map. _Res. Microbiol._ 166, 831–841. https://doi.org/10.1016/j.resmic.2015.09.009 (2015).
Article PubMed Google Scholar * Louca, S. _et al._ Function and functional redundancy in microbial systems. _Nat. Ecol. Evol._ 2, 936–943. https://doi.org/10.1038/s41559-018-0519-1
(2018). Article PubMed Google Scholar * Jousset, A. _et al._ Where less may be more: how the rare biosphere pulls ecosystems strings. _ISME J._ 11, 853–862.
https://doi.org/10.1038/ismej.2016.174 (2017). Article PubMed PubMed Central Google Scholar * Dai, T. J. _et al._ Identifying the key taxonomic categories that characterize microbial
community diversity using full-scale classification: A case study of microbial communities in the sediments of Hangzhou Bay. _FEMS Microbiol. Ecol._ https://doi.org/10.1093/femsec/fiw150
(2016). Article PubMed Google Scholar * Wilson, B. & Hayek, L. A. C. Distinguishing relative specialist and generalist species in the fossil record. _Mar. Micropa._ 119, 7–16.
https://doi.org/10.1016/j.marmicro.2015.07.001 (2015). Article Google Scholar * Wu, W. X., Logares, R., Huang, B. Q. & Hsieh, C. H. Abundant and rare picoeukaryotic sub-communities
present contrasting patterns in the epipelagic waters of marginal seas in the northwestern Pacific Ocean. _Environ. Microbiol._ 19(1), 287–300. https://doi.org/10.1111/1462-2920.13606
(2017). Article CAS PubMed Google Scholar * SD. _Soil Analysis in Agricultural Chemistry_ (China Agriculture Press, 2000). Google Scholar * Ministry of Environmental Protection, PRC.
Soil-Determination of Dry Matter and Water Content-Gravimetric Method; HJ613-2011; Ministry of Environmental Protection, PRC: Beijing, China (2011) (IN CHINESE). * Ministry of Environmental
Protection, PRC. Soil Determination of Organic Carbon-Potassium Dichromate Oxidation Spectrophotometric Method; HJ615-2011; Ministry of Environmental Protection, PRC: Beijing, China (2011)
(IN CHINESE). * Ministry of Agriculture, PRC. Soil Testing-Method for Determination of Available Phosphorus in Soil; NY/T 1121.7-2014; Ministry of Agriculture, PRC: Beijing, China (2012) (IN
CHINESE). * Ministry of Environmental Protection, PRC. Soil-Determination of Ammonium, Nitrite and Nitrate by Extraction with Potassium Chloride Solution-Spectrophotometric Methods.
HJ634-2012 Beijing, China (2012) (IN CHINESE). * Standardization Administration of China. Determination of Nitrate Nitrogen in Soil-Ultraviolet Spectrophotometry Method. GB/T 32737-2016
Beijing, China (2016) (IN CHINESE). * Bolger, A. M., Lohse, M. & Usadel,. Trimmomatic: A flexible trimmer for Illumina sequence data. _Bioinformatics_
https://doi.org/10.1093/bioinformatics/btu170 (2014). Article PubMed PubMed Central Google Scholar * Martin, M. Cut adapt removes adapter sequences from high-throughput sequencing reads.
_Embnet. J._ 17(1), 10–12. https://doi.org/10.14806/EJ.17.1.200 (2011). Article Google Scholar * Edgar, R. C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. _Nat.
Methods_ 10(10), 996–998. https://doi.org/10.1038/NMETH.2604 (2013). Article CAS PubMed Google Scholar * Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C. & Knight, R. UCHIME
improves sensitivity and speed of chimera detection. _Bioinformatics_ 27(16), 2194–2200. https://doi.org/10.1093/bioinformatics/btr381 (2011). Article CAS PubMed PubMed Central Google
Scholar * Wu, J. P. _et al._ Long-term nitrogen addition changes soil microbial community and litter decomposition rate in a subtropical forest. _Appl. Soil Ecol._ 142, 43–51.
https://doi.org/10.1016/j.apsoil.2019.05.014 (2019). Article Google Scholar * Jiao, S., Chen, W. M. & Wei, G. H. Biogeography and ecological diversity patterns of rare and abundant
bacteria in oil-contaminated soils. _Mol. Ecol._ 26(19), 5305–5317. https://doi.org/10.1111/mec.14218 (2017). Article CAS PubMed Google Scholar * Xue, Y. Y. _et al._ Distinct patterns
and processes of abundant and rare eukaryotic plankton communities following a reservoir cyanobacterial bloom. _ISME J._ 12, 2263–2277. https://doi.org/10.1038/s41396-018-0159-0 (2018).
Article CAS PubMed PubMed Central Google Scholar * Chen, W. D. _et al._ Stochastic processes shape microeukaryotic community assembly in a sub-tropical river across wet and dry seasons.
_Microbiome_ 7, 138. https://doi.org/10.1186/s40168-019-0749-8 (2019). Article PubMed PubMed Central Google Scholar * Levins, R. Evolution of diversity, efficiency and community
stability. _Am. Zool._ 10, 17–25. https://doi.org/10.1093/icb/10.1.17 (1968). Article Google Scholar * Estavillo, C., Pardini, R. & da Rocha, P. L. Forest loss and the biodiversity
threshold: An evaluation considering species habitat requirements and the use of matrix habitats. _PLoS One_ 8(12), e82369. https://doi.org/10.1371/journal.pone.0082369 (2013). Article CAS
ADS PubMed PubMed Central Google Scholar * Wu, C. _et al._ Dynamics of bacterial communities during a seasonal hypoxia at the Bohai Sea: Coupling and response between abundant and rare
populations. _J. Environ. Sci._ 111(01), 324–339. https://doi.org/10.1016/j.jes.2021.04.013 (2022). Article Google Scholar * Buee, M., Courty, P. E., Mignot, D. & Garbaye, J. Soil
niche effect on species diversity and catabolic activities in an ectomycorrhizal fungal community. _Soil Biol. Biochem._ 39, 1947–1955. https://doi.org/10.1016/j.soilbio.2007.02.016 (2007).
Article CAS Google Scholar * van der Voort, M., Kempenaar, M., van Driel, M., Raaijmakers, J. M. & Mendes, R. Impact of soil heat on reassembly of bacterial communities in the
rhizosphere microbiome and plant disease suppression. _Ecol. Lett._ 19, 375–382. https://doi.org/10.1111/ele.12567 (2016). Article PubMed Google Scholar * Xia, Z. C., Kong, C. H., Wang,
P., Chen, L. C. & Wang, S. L. Characteristics of soil microbial community structure in _Cunninghamia lanceolata_ plantation. _J. Appl. Ecol._ 23(8), 2135–2140 (2012) (IN CHINESE). CAS
Google Scholar * Liao, J. Q. _et al._ The importance of neutral and niche processes for bacterial community assembly differs between habitat generalists and specialists. _FEMS Microbiol.
Ecol._ https://doi.org/10.1093/femsec/fiw1174 (2016). Article PubMed Google Scholar * Lindh, M. V. _et al._ Local environmental conditions shape generalist but not specialist components
of microbial metacommunities in the Baltic Sea. _Front. Microbiol._ 7, 2078. https://doi.org/10.3389/fmicb.2016.02078 (2016). Article PubMed PubMed Central Google Scholar * Drury, W. H.
Rare species. _Biol. Conserv._ 6, 162–169. https://doi.org/10.1016/0006-3207(74)90061-5 (1974). Article Google Scholar * Sogin, M. L. _et al._ Microbial diversity in the deep sea and the
underexplored “rare biosphere”. _Proc. Natl. Acad. Sci. U. S. A._ 103, 12115–12120. https://doi.org/10.1073/pnas.0605127103 (2006). Article CAS ADS PubMed PubMed Central Google Scholar
* Kang, P. _et al._ A comparison of microbial composition under three tree ecosystems using the stochastic process and network complexity approaches. _Front. Microbiol._ 13, 1018077.
https://doi.org/10.3389/fmicb.2022.1018077 (2022). Article PubMed PubMed Central Google Scholar * Hearn, A. J. _et al._ Spatio-temporal ecology of sympatric felids on Borneo. Evidence
for resource partitioning?. _PLoS One_ 13(7), e0200828. https://doi.org/10.1371/journal.pone.0200828 (2018). Article CAS PubMed PubMed Central Google Scholar * Li, P. F. _et al._
Spatial variation in soil fungal communities across paddy fields in subtropical China. _mSystems_ 5(1), e00704. https://doi.org/10.1128/mSystems.00704-19 (2020). Article CAS PubMed PubMed
Central Google Scholar Download references FUNDING The authors declare no sources of funding, grants, or other support were received during the preparation of this manuscript. AUTHOR
INFORMATION AUTHORS AND AFFILIATIONS * College of Forestry Engineering, Shandong Agricultural and Engineering University, Jinan, 250100, China Zhenlu Qiu, Jie Li, Peng Wang, Dong Wang &
Jing Shu * College of Biological and Chemical Enginering, Qilu Institute of Technology, Jinan, 250200, China Li Han & Xiaojuan Gao Authors * Zhenlu Qiu View author publications You can
also search for this author inPubMed Google Scholar * Jie Li View author publications You can also search for this author inPubMed Google Scholar * Peng Wang View author publications You can
also search for this author inPubMed Google Scholar * Dong Wang View author publications You can also search for this author inPubMed Google Scholar * Li Han View author publications You
can also search for this author inPubMed Google Scholar * Xiaojuan Gao View author publications You can also search for this author inPubMed Google Scholar * Jing Shu View author
publications You can also search for this author inPubMed Google Scholar CONTRIBUTIONS Z.Q. and J.S. contributed to the study conception and design. Field survey, material collection, indoor
experiments were performed by Z.Q., J.L., P.W. and D.W. Data collection and analysis were performed by X.G. and L.H. The first draft of the manuscript was written by Z.Q. and all authors
commented on previous versions of the manuscript. All authors read and approved the fnal manuscript. CORRESPONDING AUTHOR Correspondence to Jing Shu. ETHICS DECLARATIONS COMPETING INTERESTS
The authors declare no competing interests. ADDITIONAL INFORMATION PUBLISHER'S NOTE Springer Nature remains neutral with regard to jurisdictional claims in published maps and
institutional affiliations. SUPPLEMENTARY INFORMATION SUPPLEMENTARY INFORMATION. RIGHTS AND PERMISSIONS OPEN ACCESS This article is licensed under a Creative Commons Attribution 4.0
International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the
source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's
Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not
permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit
http://creativecommons.org/licenses/by/4.0/. Reprints and permissions ABOUT THIS ARTICLE CITE THIS ARTICLE Qiu, Z., Li, J., Wang, P. _et al._ Response of soil bacteria on
habitat-specialization and abundance gradient to different afforestation types. _Sci Rep_ 13, 18181 (2023). https://doi.org/10.1038/s41598-023-44468-x Download citation * Received: 25 May
2023 * Accepted: 09 October 2023 * Published: 24 October 2023 * DOI: https://doi.org/10.1038/s41598-023-44468-x SHARE THIS ARTICLE Anyone you share the following link with will be able to
read this content: Get shareable link Sorry, a shareable link is not currently available for this article. Copy to clipboard Provided by the Springer Nature SharedIt content-sharing
initiative